

Representation of internal speech by single neurons in human supramarginal gyrus
- PMID: 38740984
- PMCID: PMC11199147
- DOI: 10.1038/s41562-024-01867-y
Representation of internal speech by single neurons in human supramarginal gyrus
Abstract
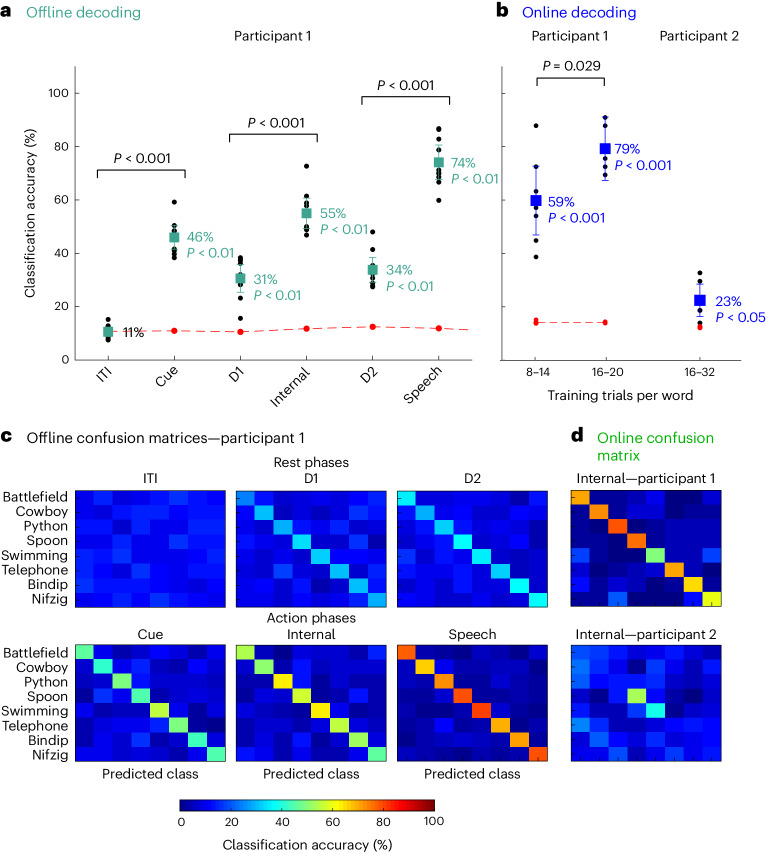
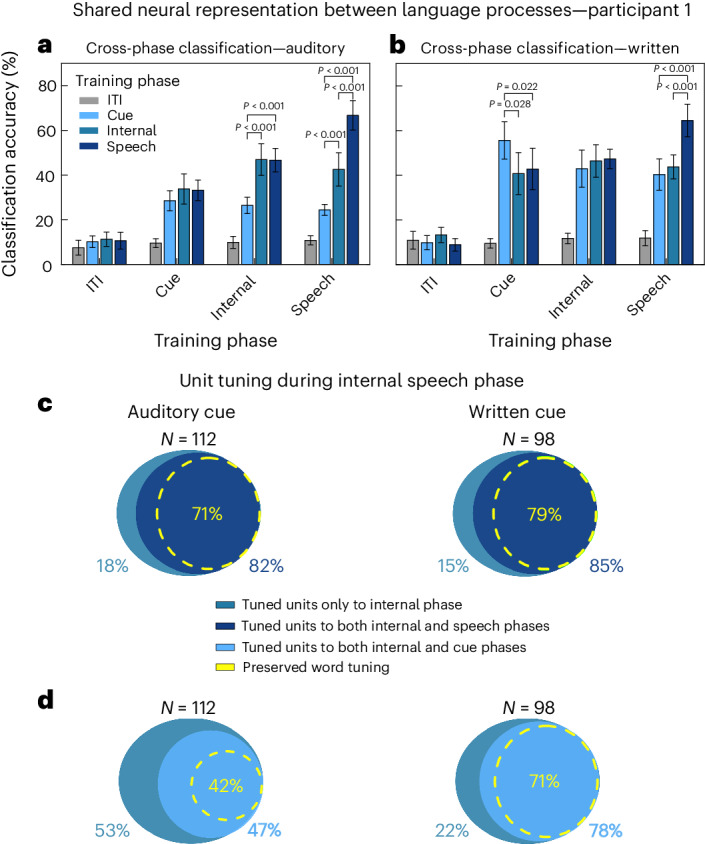
Speech brain-machine interfaces (BMIs) translate brain signals into words or audio outputs, enabling communication for people having lost their speech abilities due to diseases or injury. While important advances in vocalized, attempted and mimed speech decoding have been achieved, results for internal speech decoding are sparse and have yet to achieve high functionality. Notably, it is still unclear from which brain areas internal speech can be decoded. Here two participants with tetraplegia with implanted microelectrode arrays located in the supramarginal gyrus (SMG) and primary somatosensory cortex (S1) performed internal and vocalized speech of six words and two pseudowords. In both participants, we found significant neural representation of internal and vocalized speech, at the single neuron and population level in the SMG. From recorded population activity in the SMG, the internally spoken and vocalized words were significantly decodable. In an offline analysis, we achieved average decoding accuracies of 55% and 24% for each participant, respectively (chance level 12.5%), and during an online internal speech BMI task, we averaged 79% and 23% accuracy, respectively. Evidence of shared neural representations between internal speech, word reading and vocalized speech processes was found in participant 1. SMG represented words as well as pseudowords, providing evidence for phonetic encoding. Furthermore, our decoder achieved high classification with multiple internal speech strategies (auditory imagination/visual imagination). Activity in S1 was modulated by vocalized but not internal speech in both participants, suggesting no articulator movements of the vocal tract occurred during internal speech production. This work represents a proof-of-concept for a high-performance internal speech BMI.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
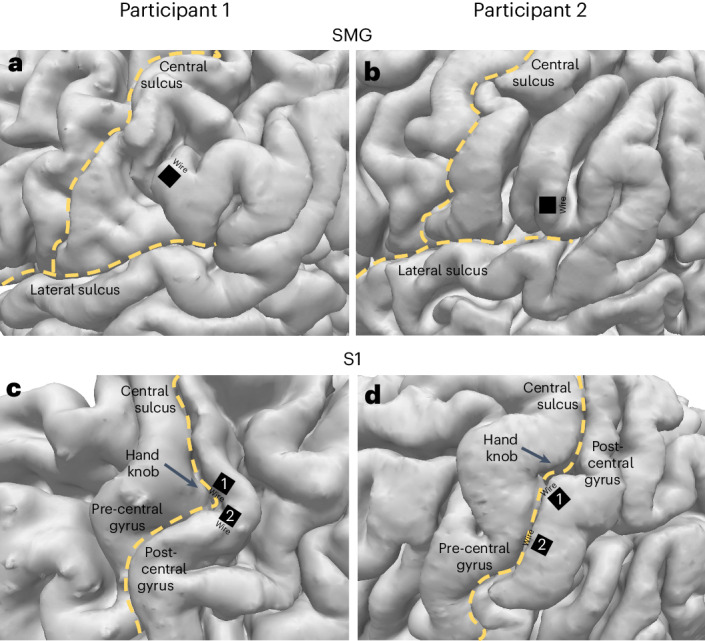
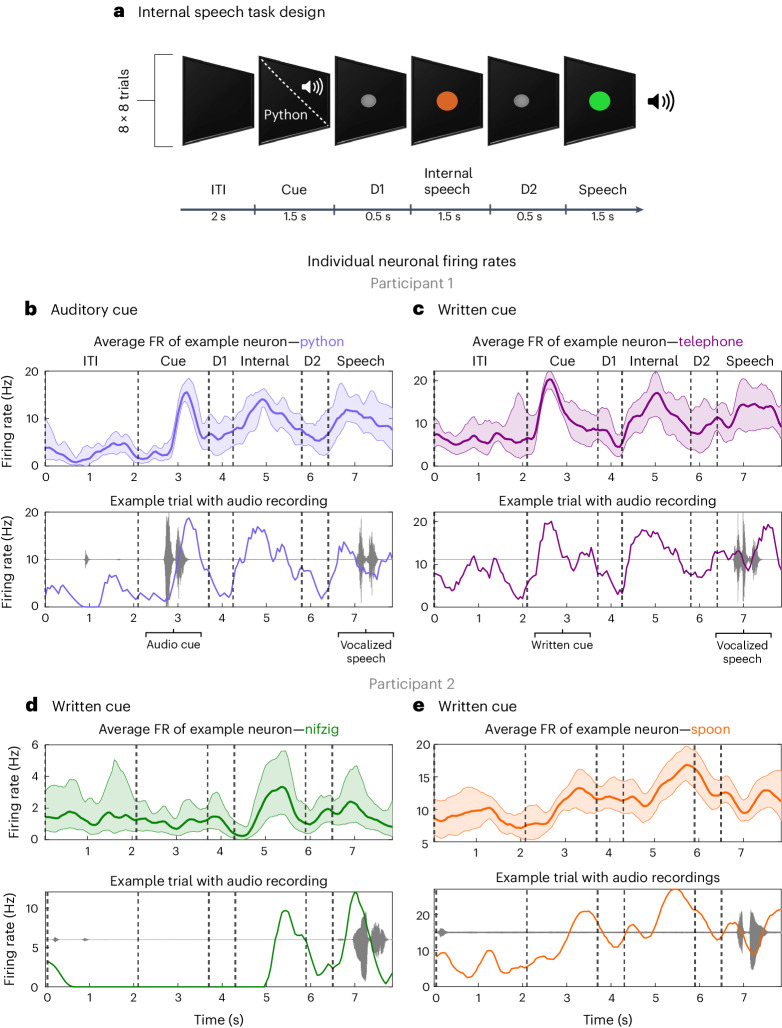
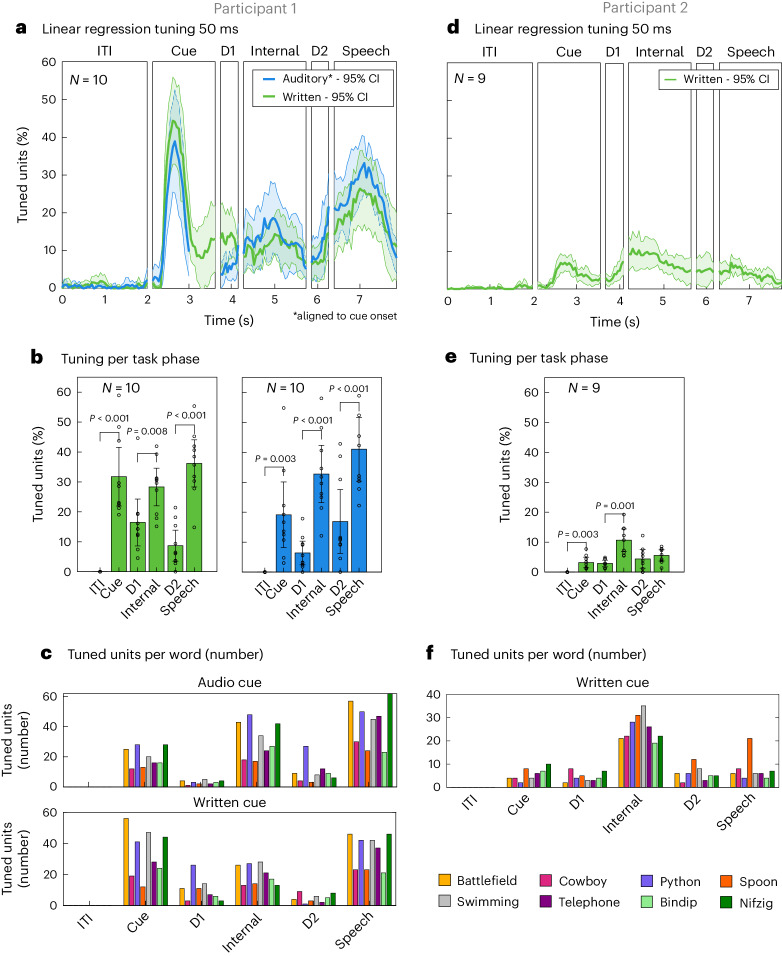
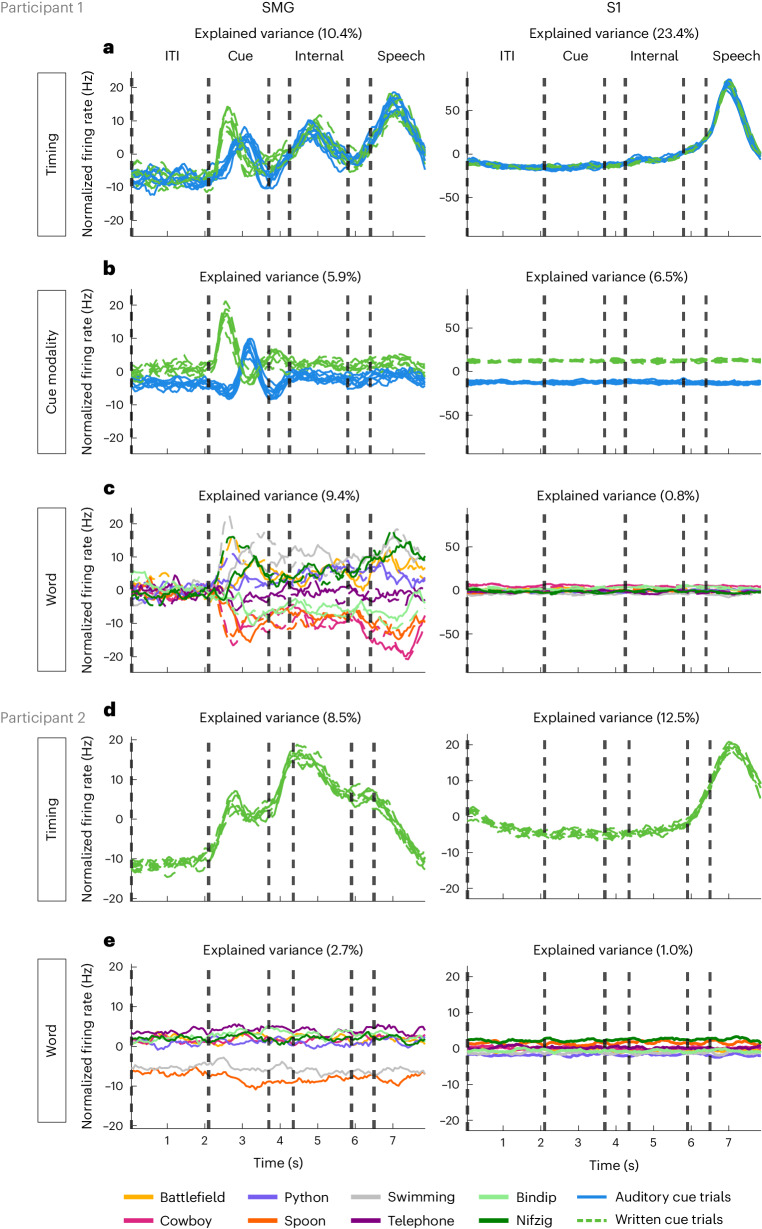
Similar articles
-
Decoding grasp and speech signals from the cortical grasp circuit in a tetraplegic human.Neuron. 2022 Jun 1;110(11):1777-1787.e3. doi: 10.1016/j.neuron.2022.03.009. Epub 2022 Mar 31. Neuron. 2022. PMID: 35364014 Free PMC article.
-
Getting a grasp on BMIs: Decoding prehension and speech signals.Neuron. 2022 Jun 1;110(11):1743-1745. doi: 10.1016/j.neuron.2022.05.004. Neuron. 2022. PMID: 35654019
-
Decoding spoken English from intracortical electrode arrays in dorsal precentral gyrus.J Neural Eng. 2020 Nov 25;17(6):066007. doi: 10.1088/1741-2552/abbfef. J Neural Eng. 2020. PMID: 33236720 Free PMC article.
-
Cognitive-motor brain-machine interfaces.J Physiol Paris. 2014 Feb;108(1):38-44. doi: 10.1016/j.jphysparis.2013.05.005. Epub 2013 Jun 15. J Physiol Paris. 2014. PMID: 23774120 Free PMC article. Review.
-
The speech neuroprosthesis.Nat Rev Neurosci. 2024 Jul;25(7):473-492. doi: 10.1038/s41583-024-00819-9. Epub 2024 May 14. Nat Rev Neurosci. 2024. PMID: 38745103 Free PMC article. Review.
Cited by
-
Whole-brain dynamics of articulatory, acoustic and semantic speech representations.Commun Biol. 2025 Mar 13;8(1):432. doi: 10.1038/s42003-025-07862-x. Commun Biol. 2025. PMID: 40082683 Free PMC article.
-
Inner speech in motor cortex and implications for speech neuroprostheses.Cell. 2025 Jul 9:S0092-8674(25)00681-6. doi: 10.1016/j.cell.2025.06.015. Online ahead of print. Cell. 2025. PMID: 40816265 Free PMC article.
-
A mind-reading brain implant that comes with password protection.Nature. 2025 Aug 14. doi: 10.1038/d41586-025-02589-5. Online ahead of print. Nature. 2025. PMID: 40813479 No abstract available.
-
Learning to operate an imagined speech Brain-Computer Interface involves the spatial and frequency tuning of neural activity.Commun Biol. 2025 Feb 20;8(1):271. doi: 10.1038/s42003-025-07464-7. Commun Biol. 2025. PMID: 39979463 Free PMC article.
-
Regional specialization of movement encoding across the primate sensorimotor cortex.Nat Commun. 2025 Jul 1;16(1):5729. doi: 10.1038/s41467-025-61172-8. Nat Commun. 2025. PMID: 40595716 Free PMC article.
References
-
- Hecht M, et al. Subjective experience and coping in ALS. Amyotroph. Lateral Scler. Other Mot. Neuron Disord. 2002;3:225–231. - PubMed
-
- Andersen, R. A. Machines that translate wants into actions. Scientific Americanhttps://www.scientificamerican.com/article/machines-that-translate-wants... (2019).
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
