

Age-related dysregulation of the retinal transcriptome in African turquoise killifish
- PMID: 38742929
- PMCID: PMC11320354
- DOI: 10.1111/acel.14192
Age-related dysregulation of the retinal transcriptome in African turquoise killifish
Abstract
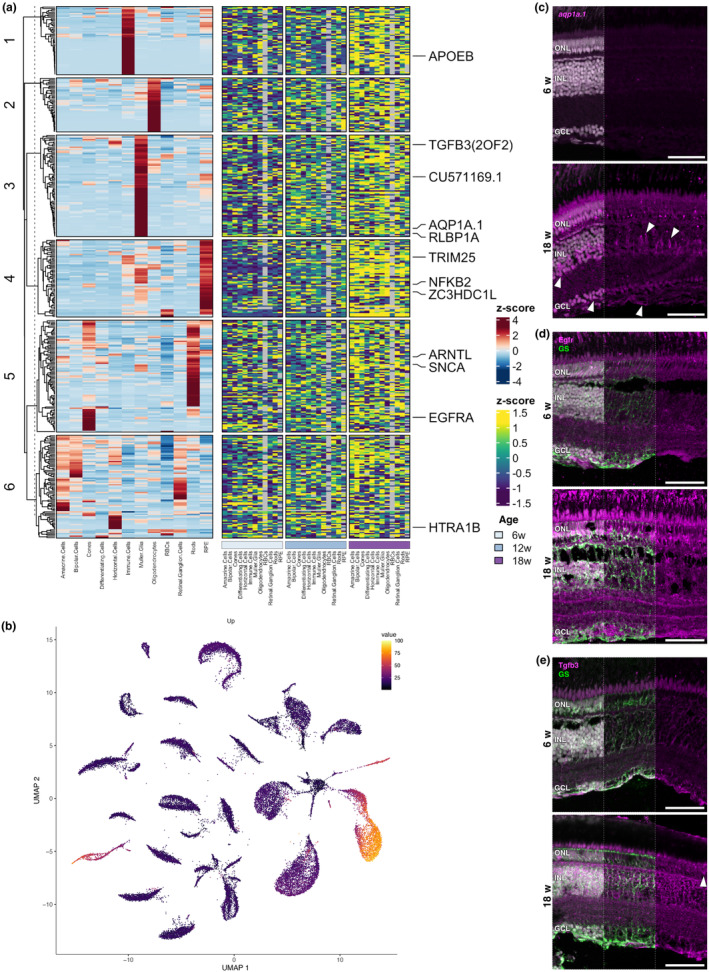
Age-related vision loss caused by retinal neurodegenerative pathologies is becoming more prevalent in our ageing society. To understand the physiological and molecular impact of ageing on retinal homeostasis, we used the short-lived African turquoise killifish, a model known to naturally develop central nervous system (CNS) ageing hallmarks and vision loss. Bulk and single-cell RNA-sequencing (scRNAseq) of three age groups (6-, 12-, and 18-week-old) identified transcriptional ageing fingerprints in the killifish retina, unveiling pathways also identified in the aged brain, including oxidative stress, gliosis, and inflammageing. These findings were comparable to observations in the ageing mouse retina. Additionally, transcriptional changes in genes related to retinal diseases, such as glaucoma and age-related macular degeneration, were observed. The cellular heterogeneity in the killifish retina was characterized, confirming the presence of all typical vertebrate retinal cell types. Data integration from age-matched samples between the bulk and scRNAseq experiments revealed a loss of cellular specificity in gene expression upon ageing, suggesting potential disruption in transcriptional homeostasis. Differential expression analysis within the identified cell types highlighted the role of glial/immune cells as important stress regulators during ageing. Our work emphasizes the value of the fast-ageing killifish in elucidating molecular signatures in age-associated retinal disease and vision decline. This study contributes to the understanding of how age-related changes in molecular pathways may impact CNS health, providing insights that may inform future therapeutic strategies for age-related pathologies.
Keywords: Nothobranchius furzeri; ageing; gliosis; inflammageing; neurodegeneration; oxidative stress; retina; transcriptomics.
© 2024 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have read and approved the manuscript and have no conflicts of interest.
Figures




Update of
-
Age-related dysregulation of the retinal transcriptome in African turquoise killifish.bioRxiv [Preprint]. 2024 Mar 11:2024.02.21.581372. doi: 10.1101/2024.02.21.581372. bioRxiv. 2024. Update in: Aging Cell. 2024 Aug;23(8):e14192. doi: 10.1111/acel.14192. PMID: 38559206 Free PMC article. Updated. Preprint.
References
-
- Ahmed, Z. M. , Riazuddin, S. , Bernstein, S. L. , Ahmed, Z. , Khan, S. , Griffith, A. J. , Morell, R. J. , Friedman, T. B. , Riazuddin, S. , & Wilcox, E. R. (2001). Mutations of the protocadherin gene PCDH15 cause usher syndrome type 1F. American Journal of Human Genetics, 69, 25–34. 10.1086/321277 - DOI - PMC - PubMed
-
- Aleksander, S. A. , Balhoff, J. , Carbon, S. , Cherry, J. M. , Drabkin, H. J. , Ebert, D. , Feuermann, M. , Gaudet, P. , Harris, N. L. , Hill, D. P. , Lee, R. , Mi, H. , Moxon, S. , Mungall, C. J. , Muruganugan, A. , Mushayahama, T. , Sternberg, P. W. , Thomas, P. D. , Van Auken, K. , et al. (2023). The gene ontology knowledgebase in 2023. Genetics, 224, iyad031. 10.1093/genetics/iyad031 - DOI - PMC - PubMed
-
- Ashburner, M. , Ball, C. A. , Blake, J. A. , Botstein, D. , Butler, H. , Cherry, J. M. , Davis, A. P. , Dolinski, K. , Dwight, S. S. , Eppig, J. T. , Harris, M. A. , Hill, D. P. , Issel‐Tarver, L. , Kasarskis, A. , Lewis, S. , Matese, J. C. , Richardson, J. E. , Ringwald, M. , Rubin, G. M. , & Sherlock, G. (2000). Gene ontology: Tool for the unification of biology. Nature Genetics, 25, 25–29. 10.1038/75556 - DOI - PMC - PubMed
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
