

Evolution of Chromosomal Inversions across an Avian Radiation
- PMID: 38743589
- PMCID: PMC11152452
- DOI: 10.1093/molbev/msae092
Evolution of Chromosomal Inversions across an Avian Radiation
Erratum in
-
Correction to: Evolution of Chromosomal Inversions across an Avian Radiation.Mol Biol Evol. 2025 Feb 3;42(2):msaf018. doi: 10.1093/molbev/msaf018. Mol Biol Evol. 2025. PMID: 39913670 Free PMC article. No abstract available.
Abstract
Chromosomal inversions are structural mutations that can play a prominent role in adaptation and speciation. Inversions segregating across species boundaries (trans-species inversions) are often taken as evidence for ancient balancing selection or adaptive introgression, but can also be due to incomplete lineage sorting. Using whole-genome resequencing data from 18 populations of 11 recognized munia species in the genus Lonchura (N = 176 individuals), we identify four large para- and pericentric inversions ranging in size from 4 to 20 Mb. All four inversions cosegregate across multiple species and predate the numerous speciation events associated with the rapid radiation of this clade across the prehistoric Sahul (Australia, New Guinea) and Bismarck Archipelago. Using coalescent theory, we infer that trans-specificity is improbable for neutrally segregating variation despite substantial incomplete lineage sorting characterizing this young radiation. Instead, the maintenance of all three autosomal inversions (chr1, chr5, and chr6) is best explained by selection acting along ecogeographic clines not observed for the collinear parts of the genome. In addition, the sex chromosome inversion largely aligns with species boundaries and shows signatures of repeated positive selection for both alleles. This study provides evidence for trans-species inversion polymorphisms involved in both adaptation and speciation. It further highlights the importance of informing selection inference using a null model of neutral evolution derived from the collinear part of the genome.
Keywords: Lonchura; ecological selection; munia; speciation; trans-species polymorphism.
© The Author(s) 2024. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures

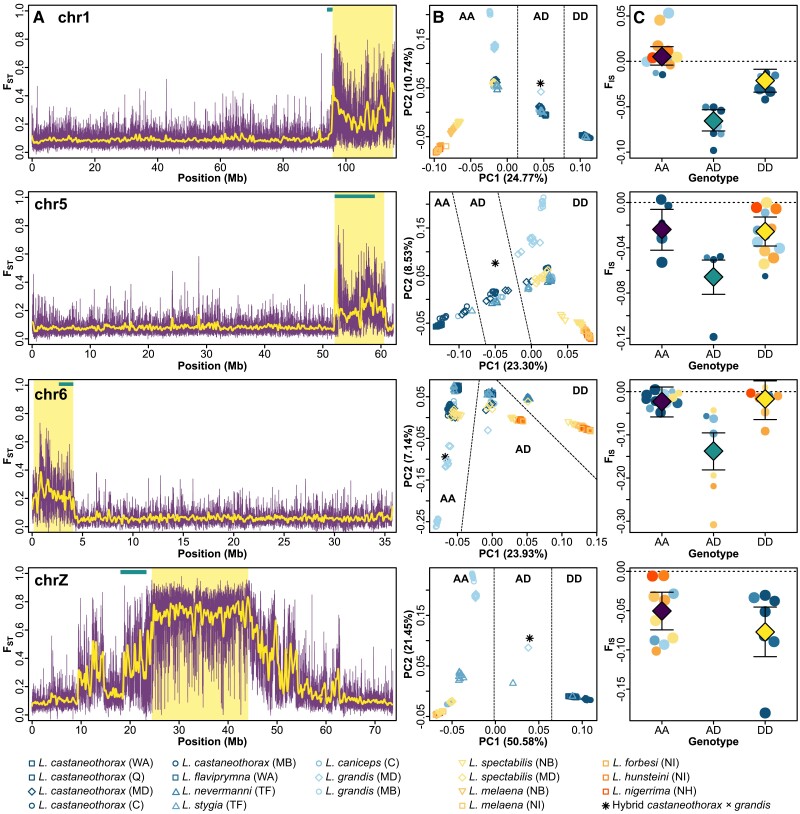
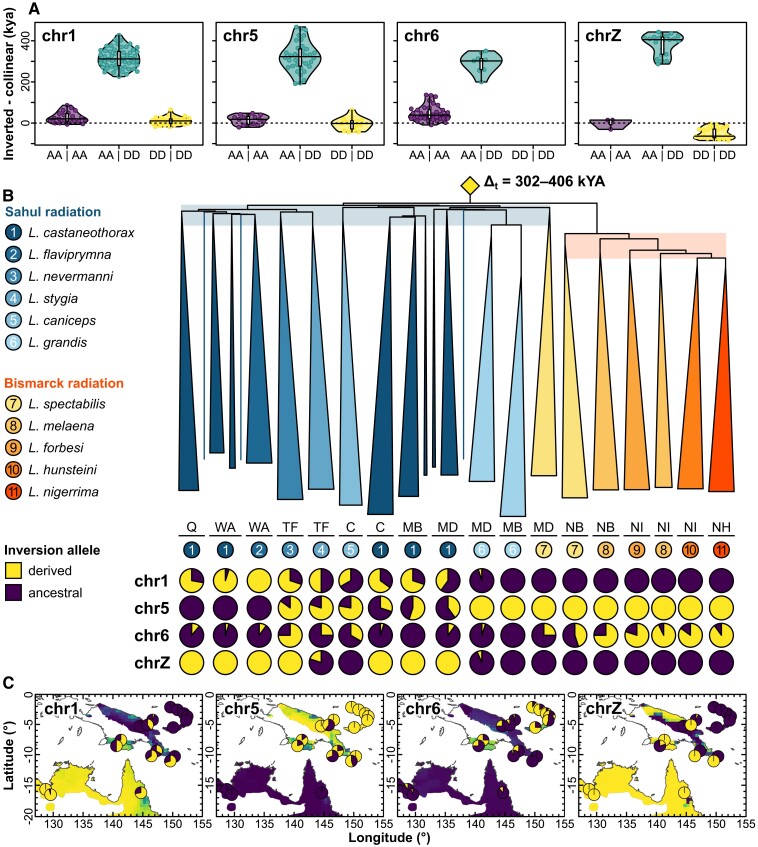


References
-
- Anderson SC, Ward EJ, English PA, Barnett LAK. sdmTMB: an R package for fast, flexible, and user-friendly generalized linear mixed effects models with spatial and spatiotemporal random fields. 2022. bioRxiv 2022.03.24.485545. 10.1101/2022.03.24.485545. preprint: not peer reviewed. - DOI
-
- Andrews S. FastQC: a quality control tool for high throughput sequence data. 2010. https://www.bioinformatics.babraham.ac.uk/projects/fastqc/.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
