

This is a preprint.
Peroxisome proliferator-activated receptor alpha is essential factor in enhanced macrophage immune function induced by angiotensin converting enzyme
- PMID: 38746124
- PMCID: PMC11092867
- DOI: 10.21203/rs.3.rs-4255086/v1
Peroxisome proliferator-activated receptor alpha is essential factor in enhanced macrophage immune function induced by angiotensin converting enzyme
Update in
-
Peroxisome proliferator-activated receptor alpha is an essential factor in enhanced macrophage immune function induced by angiotensin-converting enzyme.Cell Mol Immunol. 2025 Mar;22(3):243-259. doi: 10.1038/s41423-025-01257-y. Epub 2025 Feb 5. Cell Mol Immunol. 2025. PMID: 39910334 Free PMC article.
Abstract
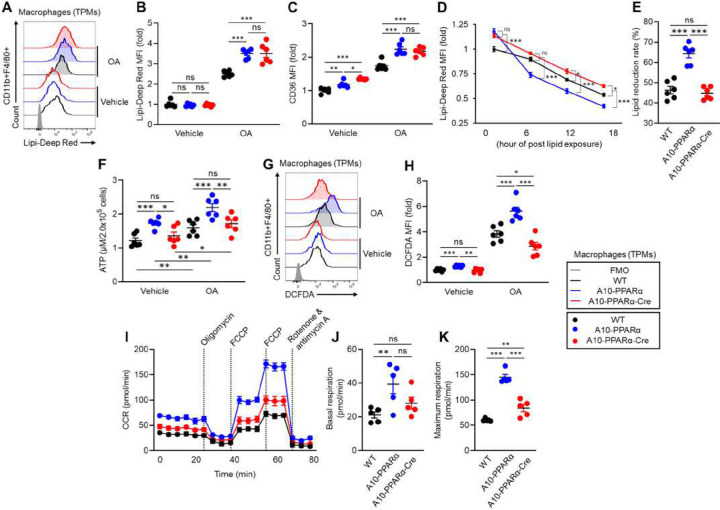
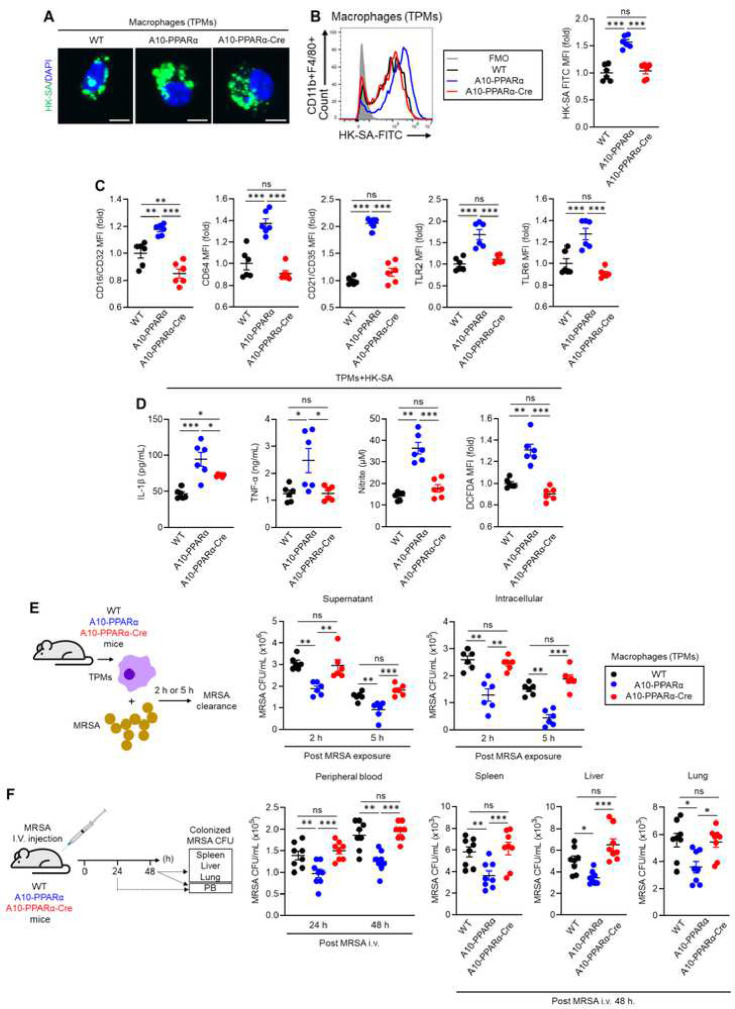
An upregulation of angiotensin-converting enzyme (ACE) expression strengthens the immune activity of myeloid lineage cells as a natural functional regulation mechanism in our immunity. ACE10/10 mice, possessing increased ACE expression in macrophages, exhibit enhanced anti-tumor immunity and anti-bactericidal effects compared to those of wild type (WT) mice, while the detailed molecular mechanism has not been elucidated yet. In this report, we demonstrate that peroxisome proliferator-activated receptor alpha (PPARα) is a key molecule in the functional upregulation of macrophages induced by ACE. The expression of PPARα, a transcription factor regulating fatty acid metabolism-associated gene expressions, was upregulated in ACE-overexpressing macrophages. To pinpoint the role of PPARα in the enhanced immune function of ACE-overexpressing macrophages, we established a line with myeloid lineage-selective PPARα depletion employing the Lysozyme 2 (LysM)-Cre system based on ACE 10/10 mice (named A10-PPARα-Cre). Interestingly, A10-PPARα-Cre mice exhibited larger B16-F10-originated tumors than original ACE 10/10 mice. PPARα depletion impaired cytokine production and antigen-presenting activity in ACE-overexpressing macrophages, resulting in reduced tumor antigen-specific CD8+ T cell activity. Additionally, the anti-bactericidal effect was also impaired in A10-PPARα-Cre mice, resulting in similar bacterial colonization to WT mice in Methicillin-Resistant Staphylococcus aureus (MRSA) infection. PPARα depletion downregulated phagocytic activity and bacteria killing in ACE-overexpressing macrophages. Moreover, THP-1-ACE-derived macrophages, as a human model, expressing upregulated PPARα exhibited enhanced cytotoxicity against B16-F10 cells and MRSA killing. These activities were further enhanced by the PPARα agonist, WY 14643, while abolished by the antagonist, GW6471, in THP-1-ACE cells. Thus, PPARα is an indispensable molecule in ACE-dependent functional upregulation of macrophages in both mice and humans.
Keywords: PPARα; angiotensin converting enzyme; anti-tumor immunity; bacterial clearance; macrophages.
Conflict of interest statement
Conflict of Interest The authors have declared that no conflict of interest exists.
Figures





References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
