

The mechanosensory channel PIEZO1 functions upstream of angiopoietin/TIE/FOXO1 signaling in lymphatic development
- PMID: 38747287
- PMCID: PMC11093609
- DOI: 10.1172/JCI176577
The mechanosensory channel PIEZO1 functions upstream of angiopoietin/TIE/FOXO1 signaling in lymphatic development
Abstract
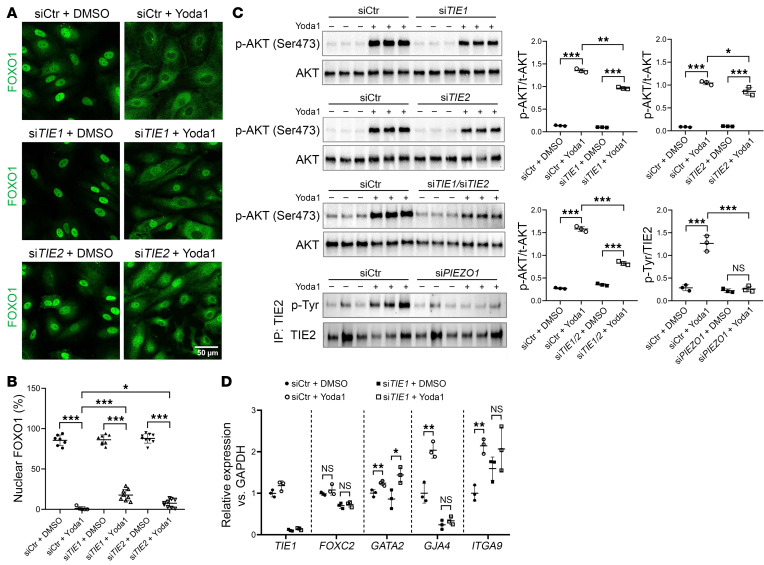
Lymphedema is a debilitating disease with no effective cure and affects an estimated 250 million individuals worldwide. Prior studies have identified mutations in piezo-type mechanosensitive ion channel component 1 (PIEZO1), angiopoietin 2 (ANGPT2), and tyrosine kinase with Ig-like and EGF-like domains 1 (TIE1) in patients with primary lymphedema. Here, we identified crosstalk between these molecules and showed that activation of the mechanosensory channel PIEZO1 in lymphatic endothelial cells (LECs) caused rapid exocytosis of the TIE ligand ANGPT2, ectodomain shedding of TIE1 by disintegrin and metalloproteinase domain-containing protein 17 (ADAM17), and increased TIE/PI3K/AKT signaling, followed by nuclear export of the transcription factor FOXO1. These data establish a functional network between lymphedema-associated genes and provide what we believe to be the first molecular mechanism bridging channel function with vascular signaling and intracellular events culminating in transcriptional regulation of genes expressed in LECs. Our study provides insights into the regulation of lymphatic function and molecular pathways involved in human disease.
Keywords: Development; Embryonic development; Endothelial cells; Ion channels; Vascular biology.
Figures







References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
