

Post-translational modification-dependent oligomerization switch in regulation of global transcription and DNA damage repair during genotoxic stress
- PMID: 38750015
- PMCID: PMC11096357
- DOI: 10.1038/s41467-024-48530-8
Post-translational modification-dependent oligomerization switch in regulation of global transcription and DNA damage repair during genotoxic stress
Abstract
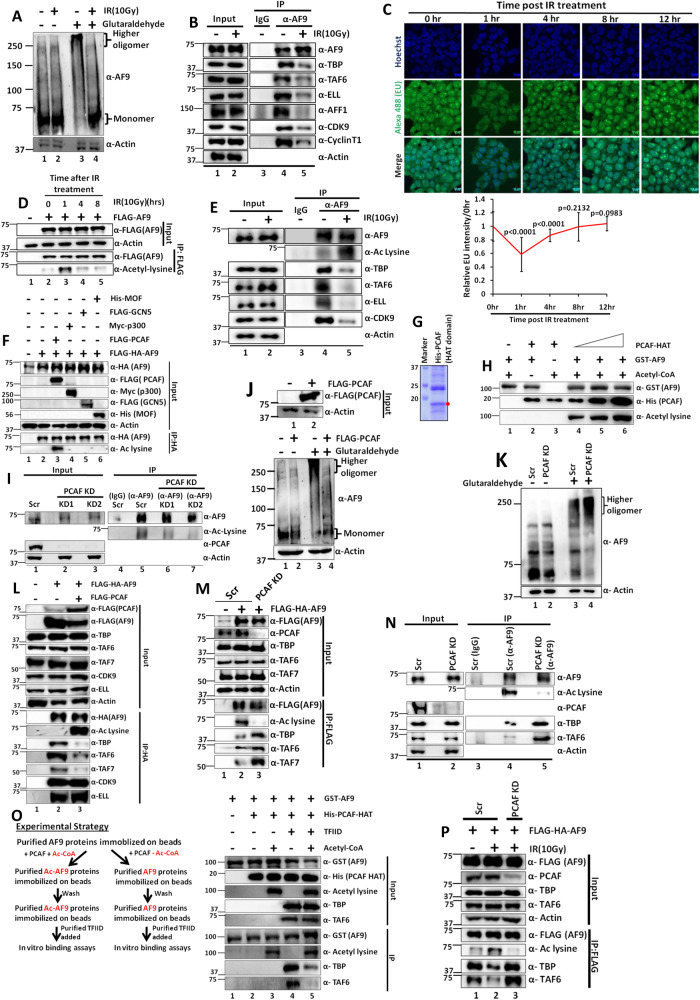
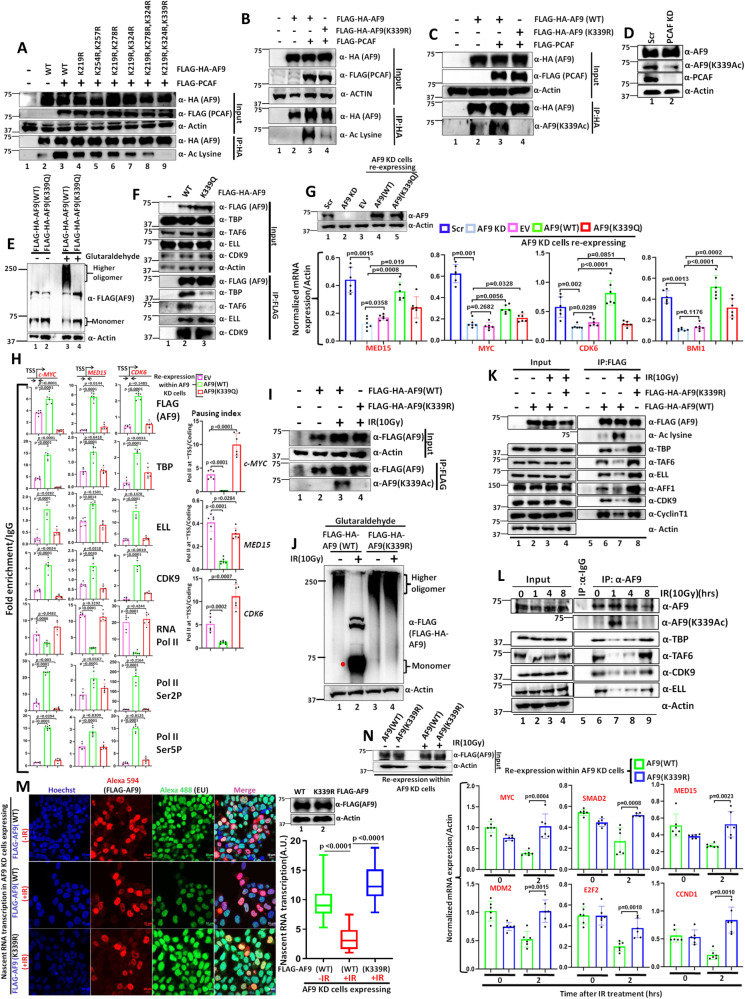
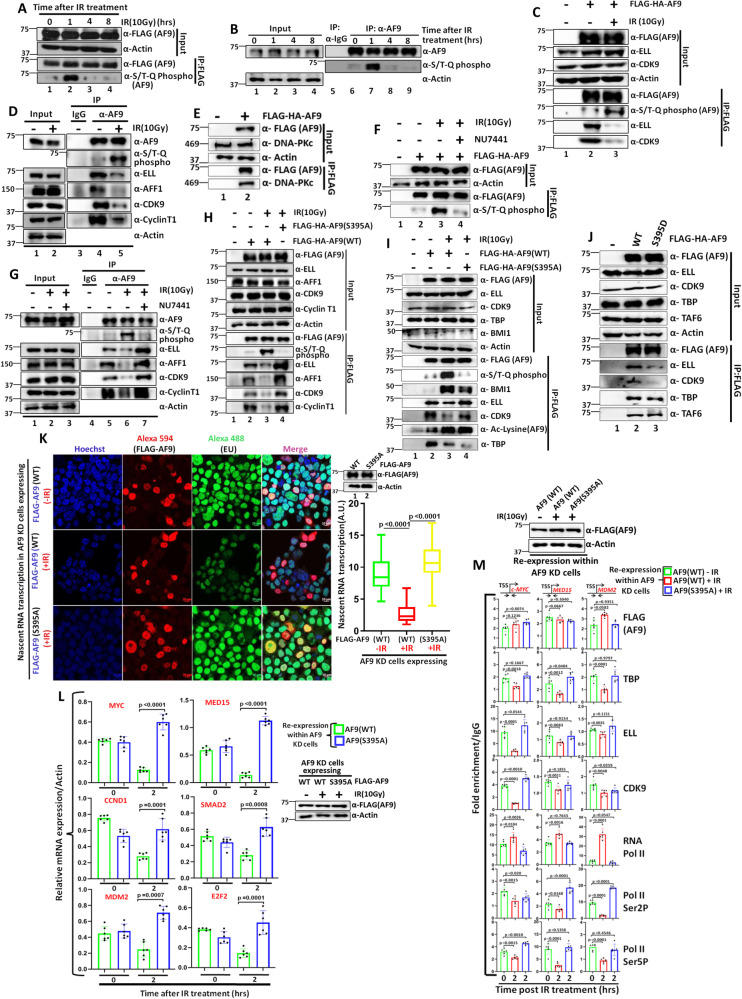
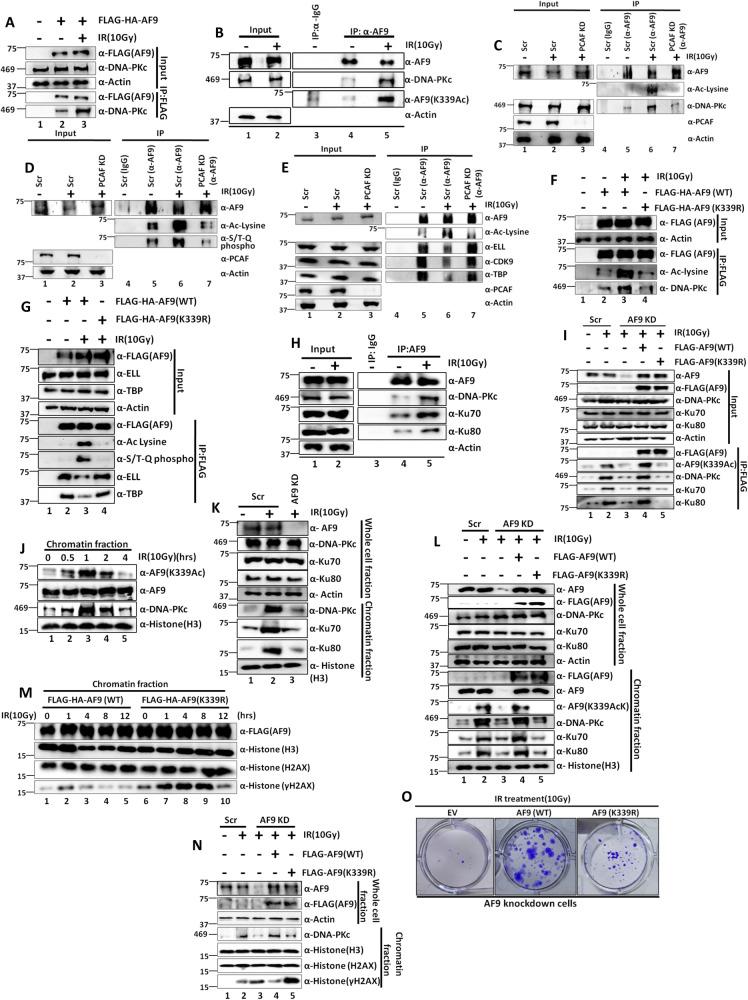
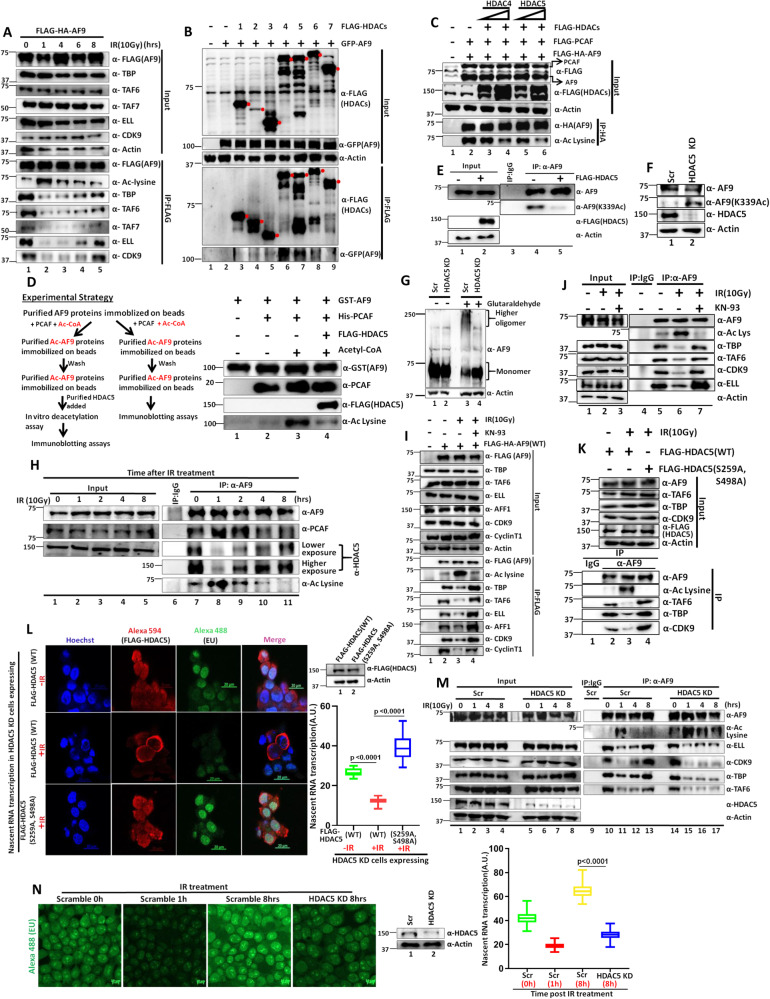
Mechanisms of functional cross-talk between global transcriptional repression and efficient DNA damage repair during genotoxic stress are poorly known. In this study, using human AF9 as representative of Super Elongation Complex (SEC) components, we delineate detailed mechanisms of these processes. Mechanistically, we describe that Poly-Serine domain-mediated oligomerization is pre-requisite for AF9 YEATS domain-mediated TFIID interaction-dependent SEC recruitment at the promoter-proximal region for release of paused RNA polymerase II. Interestingly, during genotoxic stress, CaMKII-mediated phosphorylation-dependent nuclear export of AF9-specific deacetylase HDAC5 enhances concomitant PCAF-mediated acetylation of K339 residue. This causes monomerization of AF9 and reduces TFIID interaction for transcriptional downregulation. Furthermore, the K339 acetylation-dependent enhanced AF9-DNA-PKc interaction leads to phosphorylation at S395 residue which reduces AF9-SEC interaction resulting in transcriptional downregulation and efficient repair of DNA damage. After repair, nuclear re-entry of HDAC5 reduces AF9 acetylation and restores its TFIID and SEC interaction to restart transcription.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
AFF1 acetylation by p300 temporally inhibits transcription during genotoxic stress response.Proc Natl Acad Sci U S A. 2019 Oct 29;116(44):22140-22151. doi: 10.1073/pnas.1907097116. Epub 2019 Oct 14. Proc Natl Acad Sci U S A. 2019. PMID: 31611376 Free PMC article.
-
ATM-mediated ELL phosphorylation enhances its self-association through increased EAF1 interaction and inhibits global transcription during genotoxic stress.Nucleic Acids Res. 2022 Oct 28;50(19):10995-11012. doi: 10.1093/nar/gkac943. Nucleic Acids Res. 2022. PMID: 36305813 Free PMC article.
-
Human Polymerase-Associated Factor complex (PAFc) connects the Super Elongation Complex (SEC) to RNA polymerase II on chromatin.Proc Natl Acad Sci U S A. 2011 Sep 6;108(36):E636-45. doi: 10.1073/pnas.1107107108. Epub 2011 Aug 22. Proc Natl Acad Sci U S A. 2011. PMID: 21873227 Free PMC article.
-
Collaboration of MLLT1/ENL, Polycomb and ATM for transcription and genome integrity.Nucleus. 2016 Apr 25;7(2):138-45. doi: 10.1080/19491034.2016.1177681. Nucleus. 2016. PMID: 27310306 Free PMC article. Review.
-
TRIM28 as a novel transcriptional elongation factor.BMC Mol Biol. 2015 Aug 21;16:14. doi: 10.1186/s12867-015-0040-x. BMC Mol Biol. 2015. PMID: 26293668 Free PMC article. Review.
Cited by
-
The TOP-2/condensin II axis silences transcription during germline specification in C. elegans.G3 (Bethesda). 2024 Oct 3;14(12):jkae236. doi: 10.1093/g3journal/jkae236. Online ahead of print. G3 (Bethesda). 2024. PMID: 39358855 Free PMC article.
-
LncRNA938/ TAF9/TTK axis promotes EMT and serves as a therapeutic target in hepatoblastoma.J Transl Med. 2025 Aug 21;23(1):946. doi: 10.1186/s12967-025-06809-4. J Transl Med. 2025. PMID: 40841910
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
