

Phage protein Gp11 blocks Staphylococcus aureus cell division by inhibiting peptidoglycan biosynthesis
- PMID: 38752726
- PMCID: PMC11237401
- DOI: 10.1128/mbio.00679-24
Phage protein Gp11 blocks Staphylococcus aureus cell division by inhibiting peptidoglycan biosynthesis
Abstract
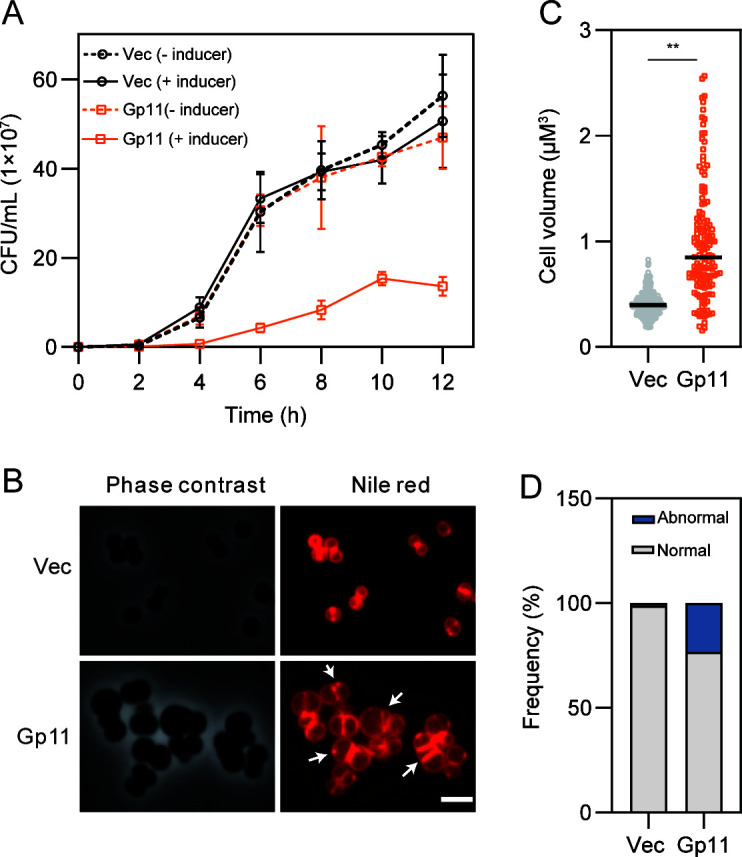
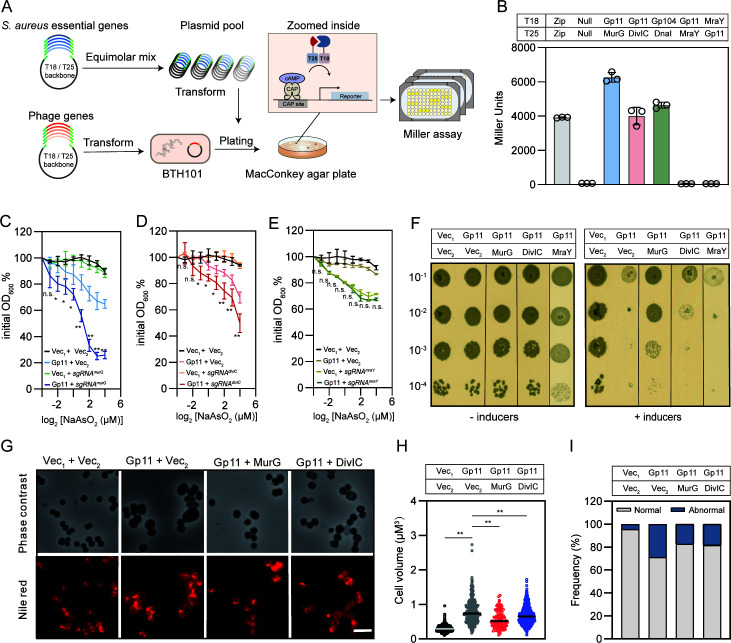
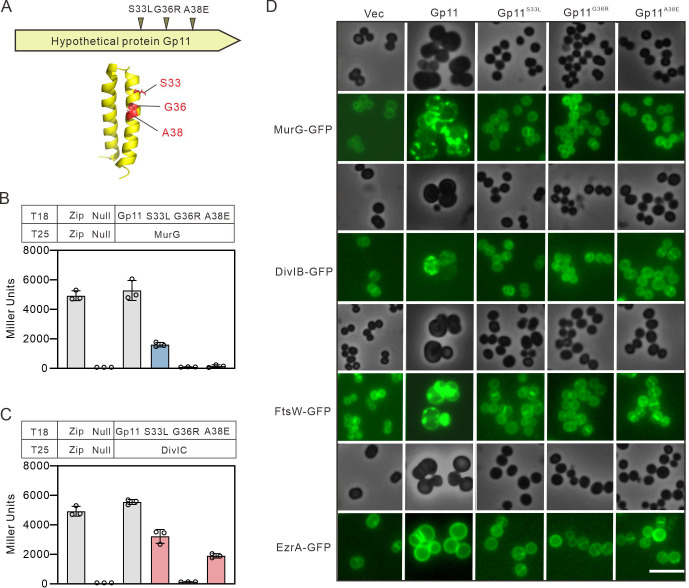
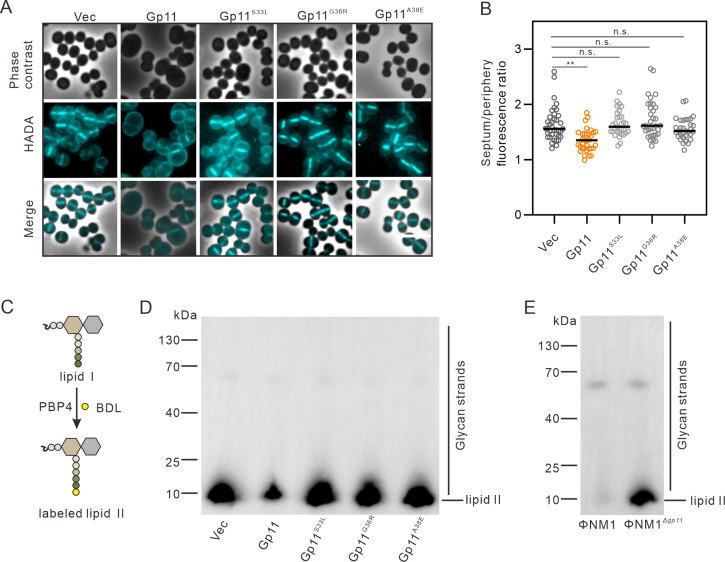
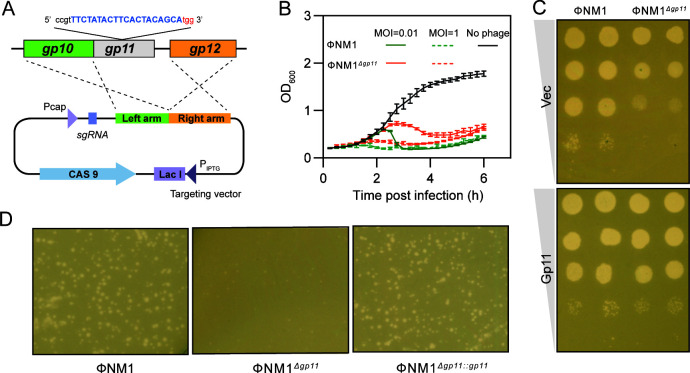
Phages and bacteria have a long history of co-evolution. However, these dynamics of phage-host interactions are still largely unknown; identification of phage inhibitors that remodel host metabolism will provide valuable information for target development for antimicrobials. Here, we perform a comprehensive screen for early-gene products of ΦNM1 that inhibit cell growth in Staphylococcus aureus. A small membrane protein, Gp11, with inhibitory effects on S. aureus cell division was identified. A bacterial two-hybrid library containing 345 essential S. aureus genes was constructed to screen for targets of Gp11, and Gp11 was found to interact with MurG and DivIC. Defects in cell growth and division caused by Gp11 were dependent on MurG and DivIC, which was further confirmed using CRISPRi hypersensitivity assay. Gp11 interacts with MurG, the protein essential for cell wall formation, by inhibiting the production of lipid II to regulate peptidoglycan (PG) biosynthesis on the cell membrane. Gp11 also interacts with cell division protein DivIC, an essential part of the division machinery necessary for septal cell wall assembly, to disrupt the recruitment of division protein FtsW. Mutations in Gp11 result in loss of its ability to cause growth defects, whereas infection with phage in which the gp11 gene has been deleted showed a significant increase in lipid II production in S. aureus. Together, our findings reveal that a phage early-gene product interacts with essential host proteins to disrupt PG biosynthesis and block S. aureus cell division, suggesting a potential pathway for the development of therapeutic approaches to treat pathogenic bacterial infections.
Importance: Understanding the interplay between phages and their hosts is important for the development of novel therapies against pathogenic bacteria. Although phages have been used to control methicillin-resistant Staphylococcus aureus infections, our knowledge related to the processes in the early stages of phage infection is still limited. Owing to the fact that most of the phage early proteins have been classified as hypothetical proteins with uncertain functions, we screened phage early-gene products that inhibit cell growth in S. aureus, and one protein, Gp11, selectively targets essential host genes to block the synthesis of the peptidoglycan component lipid II, ultimately leading to cell growth arrest in S. aureus. Our study provides a novel insight into the strategy by which Gp11 blocks essential host cellular metabolism to influence phage-host interaction. Importantly, dissecting the interactions between phages and host cells will contribute to the development of new and effective therapies to treat bacterial infections.
Keywords: Staphylococcus aureus; essential gene; lipid II; peptidoglycan; phage.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The Staphylococcus aureus cell division protein, DivIC, interacts with the cell wall and controls its biosynthesis.Commun Biol. 2022 Nov 11;5(1):1228. doi: 10.1038/s42003-022-04161-7. Commun Biol. 2022. PMID: 36369270 Free PMC article.
-
Penicillin-Binding Protein 1 (PBP1) of Staphylococcus aureus Has Multiple Essential Functions in Cell Division.mBio. 2022 Aug 30;13(4):e0066922. doi: 10.1128/mbio.00669-22. Epub 2022 Jun 15. mBio. 2022. PMID: 35703435 Free PMC article.
-
A Time-Resolved FRET Assay Identifies a Small Molecule that Inhibits the Essential Bacterial Cell Wall Polymerase FtsW.Angew Chem Int Ed Engl. 2023 Jun 19;62(25):e202301522. doi: 10.1002/anie.202301522. Epub 2023 May 11. Angew Chem Int Ed Engl. 2023. PMID: 37099323 Free PMC article.
-
Staphylococcal Protein Secretion and Envelope Assembly.Microbiol Spectr. 2019 Jul;7(4):10.1128/microbiolspec.gpp3-0070-2019. doi: 10.1128/microbiolspec.GPP3-0070-2019. Microbiol Spectr. 2019. PMID: 31267890 Free PMC article. Review.
-
Phage single-gene lysis: Finding the weak spot in the bacterial cell wall.J Biol Chem. 2019 Mar 8;294(10):3350-3358. doi: 10.1074/jbc.TM118.001773. Epub 2018 Nov 12. J Biol Chem. 2019. PMID: 30420429 Free PMC article. Review.
Cited by
-
Bacteriophage P1 protein Icd inhibits bacterial division by targeting FtsZ.Front Microbiol. 2025 Feb 26;16:1533694. doi: 10.3389/fmicb.2025.1533694. eCollection 2025. Front Microbiol. 2025. PMID: 40078545 Free PMC article.
-
Structural basis for Salmonella infection by two Microviridae phages.Commun Biol. 2025 Aug 6;8(1):1166. doi: 10.1038/s42003-025-08595-7. Commun Biol. 2025. PMID: 40770068 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
