

Peripheral preprocessing in Drosophila facilitates odor classification
- PMID: 38753511
- PMCID: PMC11126917
- DOI: 10.1073/pnas.2316799121
Peripheral preprocessing in Drosophila facilitates odor classification
Abstract
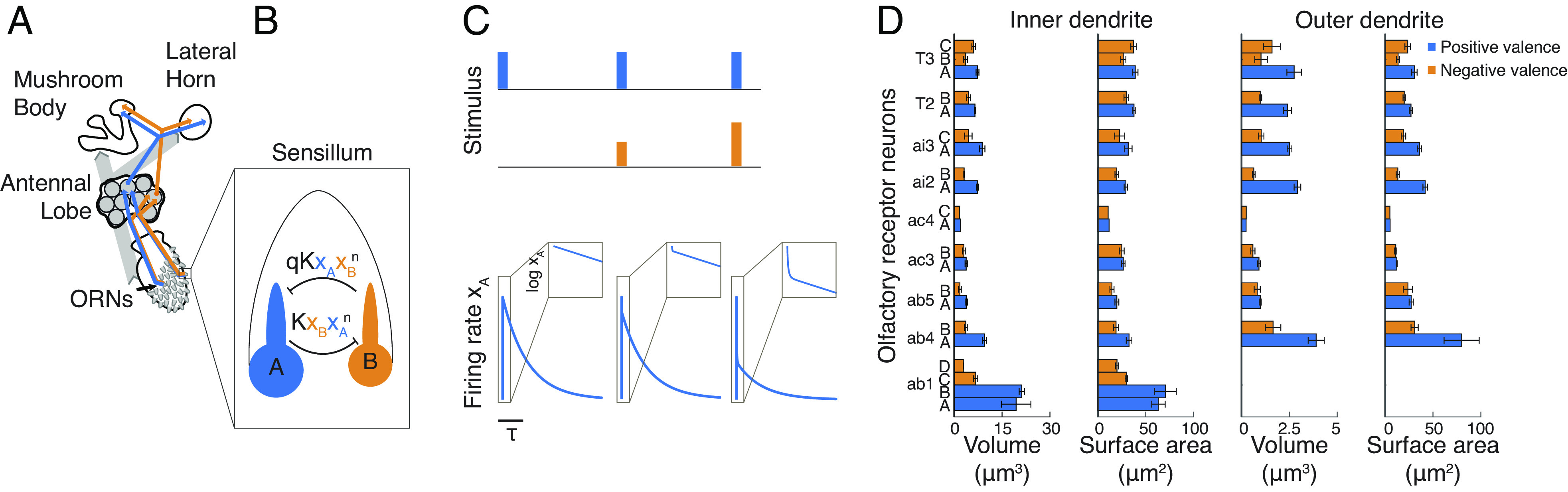
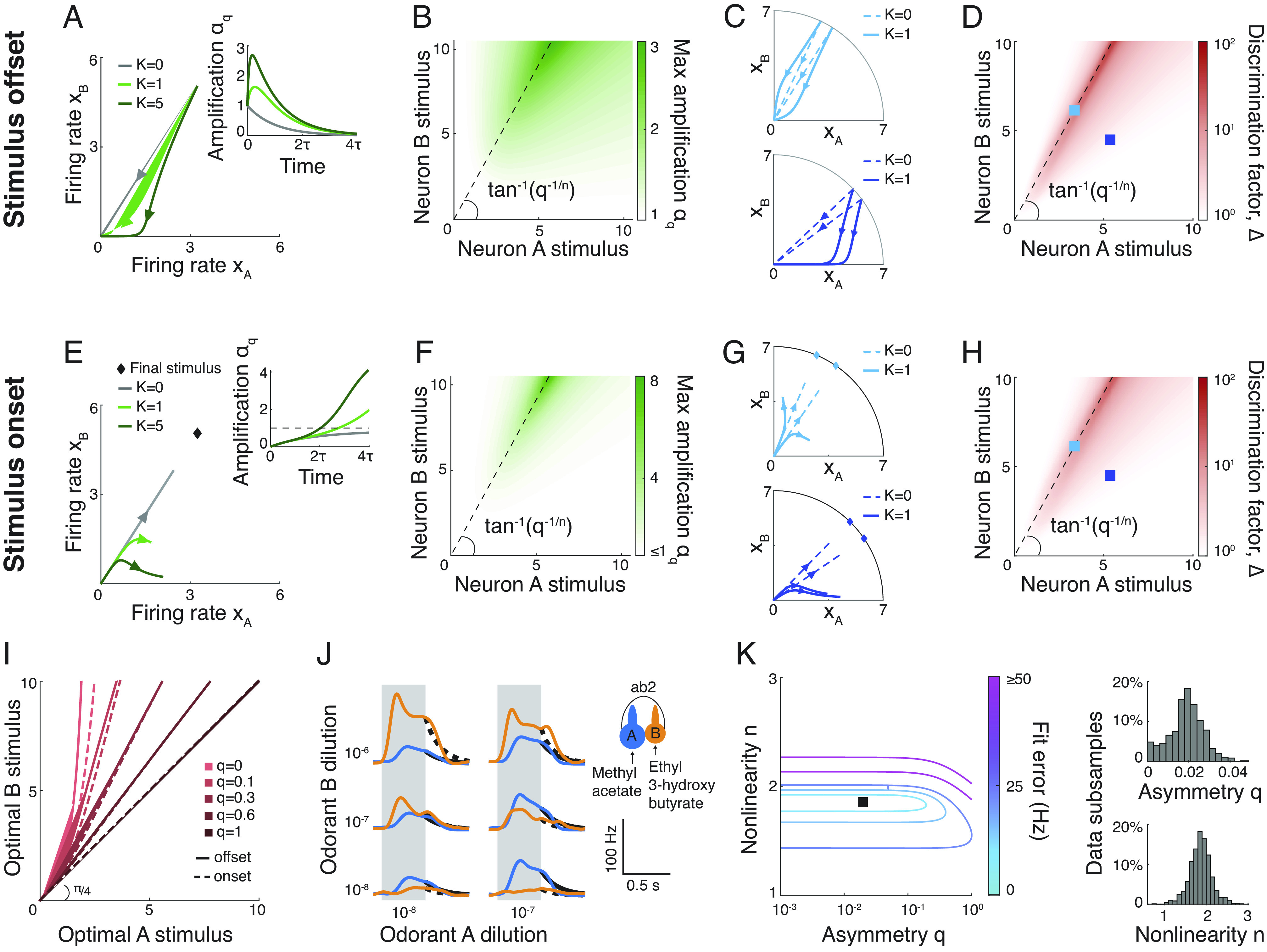
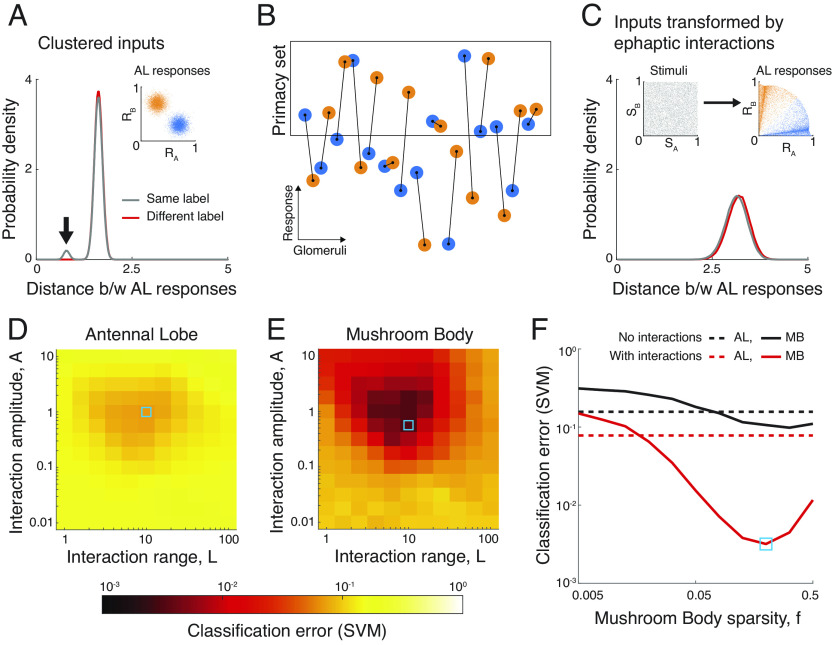
The mammalian brain implements sophisticated sensory processing algorithms along multilayered ("deep") neural networks. Strategies that insects use to meet similar computational demands, while relying on smaller nervous systems with shallow architectures, remain elusive. Using Drosophila as a model, we uncover the algorithmic role of odor preprocessing by a shallow network of compartmentalized olfactory receptor neurons. Each compartment operates as a ratiometric unit for specific odor-mixtures. This computation arises from a simple mechanism: electrical coupling between two differently sized neurons. We demonstrate that downstream synaptic connectivity is shaped to optimally leverage amplification of a hedonic value signal in the periphery. Furthermore, peripheral preprocessing is shown to markedly improve novel odor classification in a higher brain center. Together, our work highlights a far-reaching functional role of the sensory periphery for downstream processing. By elucidating the implementation of powerful computations by a shallow network, we provide insights into general principles of efficient sensory processing algorithms.
Keywords: Drosophila; connectome; olfaction; sensory periphery; shallow neural network.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures

Update of
-
Shallow networks run deep: Peripheral preprocessing facilitates odor classification.bioRxiv [Preprint]. 2023 Jul 25:2023.07.23.550211. doi: 10.1101/2023.07.23.550211. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2024 May 21;121(21):e2316799121. doi: 10.1073/pnas.2316799121. PMID: 37546820 Free PMC article. Updated. Preprint.
Similar articles
-
Shallow networks run deep: Peripheral preprocessing facilitates odor classification.bioRxiv [Preprint]. 2023 Jul 25:2023.07.23.550211. doi: 10.1101/2023.07.23.550211. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2024 May 21;121(21):e2316799121. doi: 10.1073/pnas.2316799121. PMID: 37546820 Free PMC article. Updated. Preprint.
-
Early olfactory processing in Drosophila: mechanisms and principles.Annu Rev Neurosci. 2013 Jul 8;36:217-41. doi: 10.1146/annurev-neuro-062111-150533. Annu Rev Neurosci. 2013. PMID: 23841839 Free PMC article. Review.
-
Valence opponency in peripheral olfactory processing.Proc Natl Acad Sci U S A. 2022 Feb 1;119(5):e2120134119. doi: 10.1073/pnas.2120134119. Proc Natl Acad Sci U S A. 2022. PMID: 35091473 Free PMC article.
-
Processing of odor mixtures in the Drosophila antennal lobe reveals both global inhibition and glomerulus-specific interactions.J Neurosci. 2007 Oct 31;27(44):11966-77. doi: 10.1523/JNEUROSCI.3099-07.2007. J Neurosci. 2007. PMID: 17978037 Free PMC article.
-
Odor representations in the mammalian olfactory bulb.Wiley Interdiscip Rev Syst Biol Med. 2010 Sep-Oct;2(5):603-611. doi: 10.1002/wsbm.85. Wiley Interdiscip Rev Syst Biol Med. 2010. PMID: 20836051 Review.
Cited by
-
Nonlinear high-activity neuronal excitation enhances odor discrimination.Curr Biol. 2025 Apr 7;35(7):1521-1538.e5. doi: 10.1016/j.cub.2025.02.034. Epub 2025 Mar 18. Curr Biol. 2025. PMID: 40107267 Free PMC article.
References
-
- Celani A., Villermaux E., Vergassola M., Odor landscapes in turbulent environments. Phys. Rev. X 4, 041015 (2014).
-
- Reddy G., Murthy V. N., Vergassola M., Olfactory sensing and navigation in turbulent environments. Annu. Rev. Condens. Matter Phys. 13, 191–213 (2022).
MeSH terms
Grants and funding
- R21DC020536/HHS | NIH | National Institute on Deafness and Other Communication Disorders (NIDCD)
- R01NS135853/HHS | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
- R01 NS135853/NS/NINDS NIH HHS/United States
- R21AI169343/HHS | NIH | National Institute of Allergy and Infectious Diseases (NIAID)
- R21 AI169343/AI/NIAID NIH HHS/United States
- R01DC016466/HHS | NIH | National Institute on Deafness and Other Communication Disorders (NIDCD)
- DE-SC0022042/DOE | Office of Science (SC)
- D21AP10162-00/DOD | ARPA | Biological Technologies Office (BTO)
- IRG 2022/UC | UCSD | Kavli Institute for Brain and Mind, University of California, San Diego (KIBM, UCSD)
- R21 DC020536/DC/NIDCD NIH HHS/United States
- R01 DC016466/DC/NIDCD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
