

Autophagy counters inflammation-driven glycolytic impairment in aging hematopoietic stem cells
- PMID: 38754428
- PMCID: PMC11350610
- DOI: 10.1016/j.stem.2024.04.020
Autophagy counters inflammation-driven glycolytic impairment in aging hematopoietic stem cells
Abstract
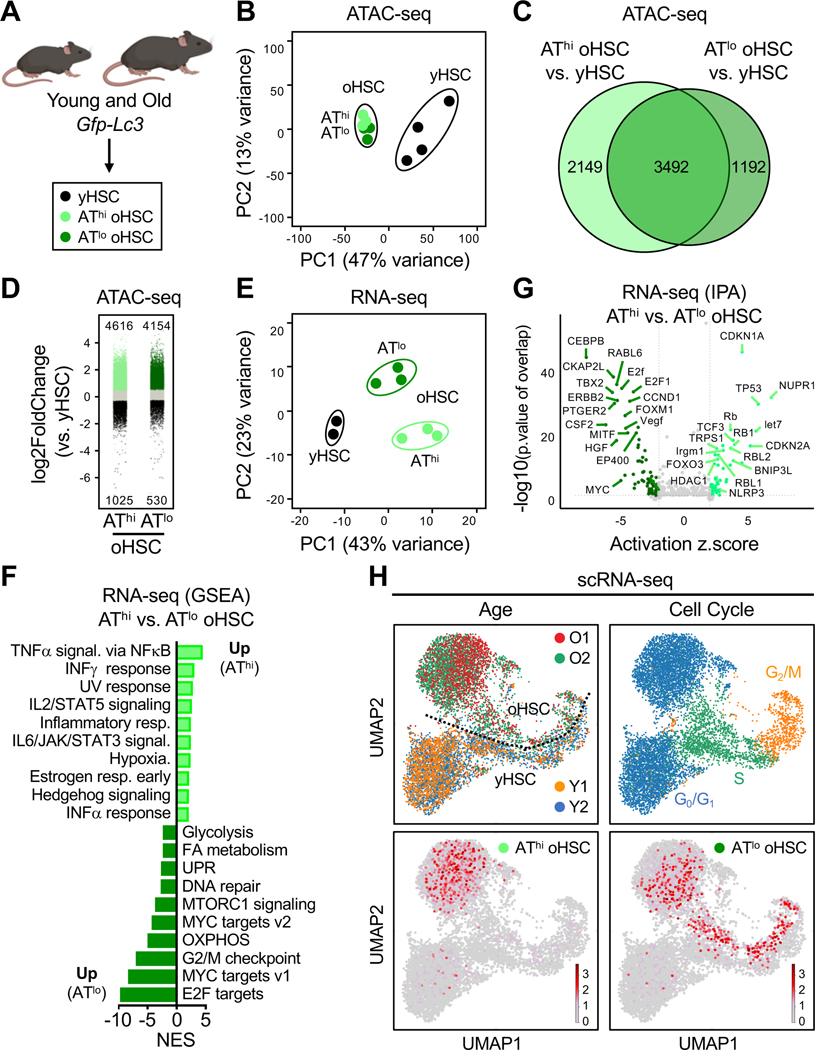
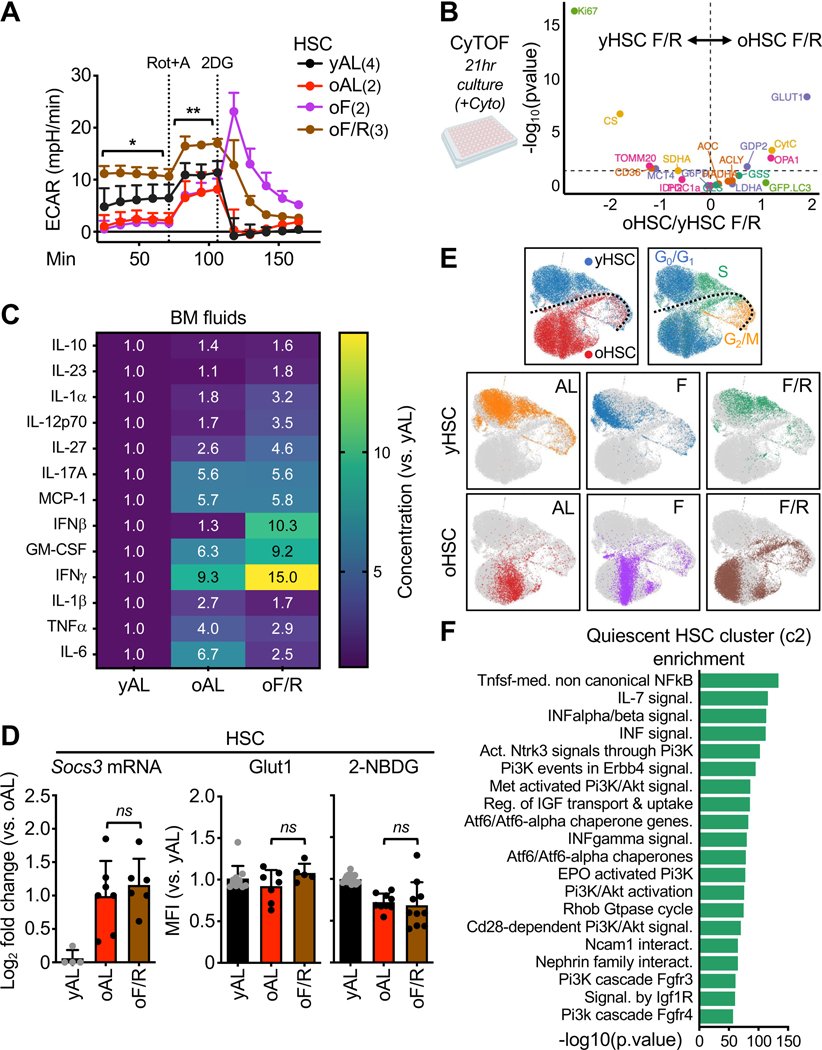
Autophagy is central to the benefits of longevity signaling programs and to hematopoietic stem cell (HSC) response to nutrient stress. With age, a subset of HSCs increases autophagy flux and preserves regenerative capacity, but the signals triggering autophagy and maintaining the functionality of autophagy-activated old HSCs (oHSCs) remain unknown. Here, we demonstrate that autophagy is an adaptive cytoprotective response to chronic inflammation in the aging murine bone marrow (BM) niche. We find that inflammation impairs glucose uptake and suppresses glycolysis in oHSCs through Socs3-mediated inhibition of AKT/FoxO-dependent signaling, with inflammation-mediated autophagy engagement preserving functional quiescence by enabling metabolic adaptation to glycolytic impairment. Moreover, we show that transient autophagy induction via a short-term fasting/refeeding paradigm normalizes glycolytic flux and significantly boosts oHSC regenerative potential. Our results identify inflammation-driven glucose hypometabolism as a key driver of HSC dysfunction with age and establish autophagy as a targetable node to reset oHSC regenerative capacity.
Keywords: aging; autophagy; hematopoietic stem cells; inflammation; metabolism; regeneration.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests E.P. is a member of the Cell Stem Cell advisory board.
Figures





Update of
-
Autophagy counters inflammation-driven glycolytic impairment in aging hematopoietic stem cells.bioRxiv [Preprint]. 2023 Aug 19:2023.08.17.553736. doi: 10.1101/2023.08.17.553736. bioRxiv. 2023. Update in: Cell Stem Cell. 2024 Jul 5;31(7):1020-1037.e9. doi: 10.1016/j.stem.2024.04.020. PMID: 37645930 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
