

Mycobacterial biotin synthases require an auxiliary protein to convert dethiobiotin into biotin
- PMID: 38755122
- PMCID: PMC11099021
- DOI: 10.1038/s41467-024-48448-1
Mycobacterial biotin synthases require an auxiliary protein to convert dethiobiotin into biotin
Abstract
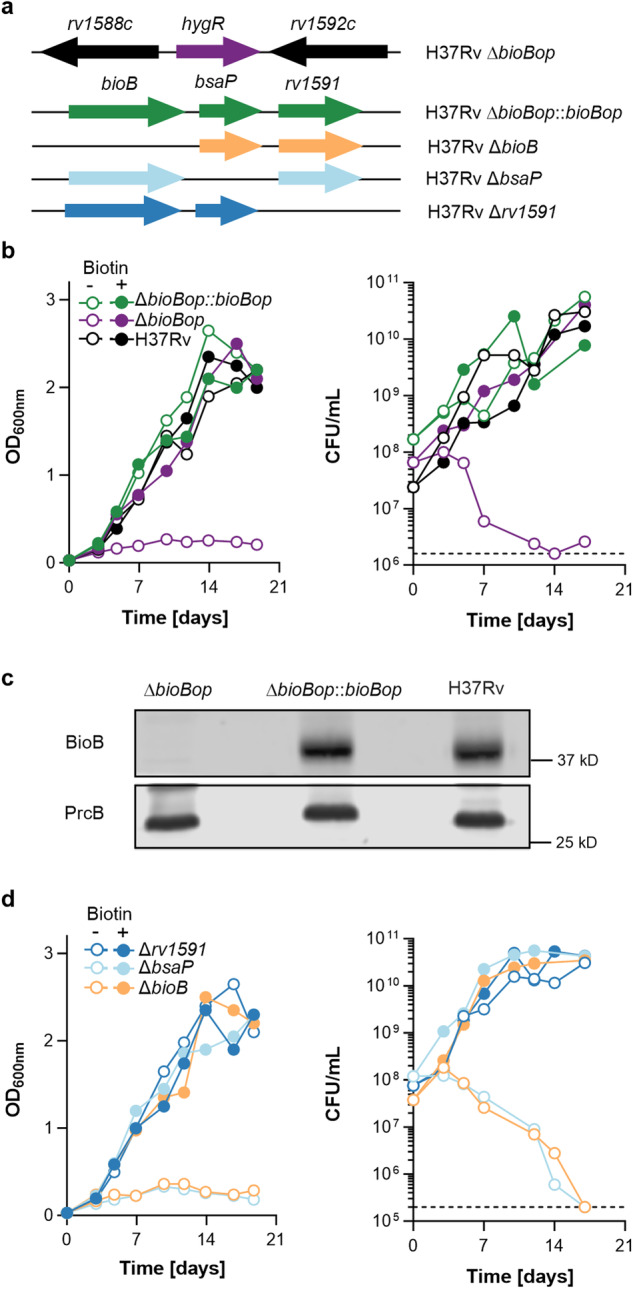
Lipid biosynthesis in the pathogen Mycobacterium tuberculosis depends on biotin for posttranslational modification of key enzymes. However, the mycobacterial biotin synthetic pathway is not fully understood. Here, we show that rv1590, a gene of previously unknown function, is required by M. tuberculosis to synthesize biotin. Chemical-generic interaction experiments mapped the function of rv1590 to the conversion of dethiobiotin to biotin, which is catalyzed by biotin synthases (BioB). Biochemical studies confirmed that in contrast to BioB of Escherichia coli, BioB of M. tuberculosis requires Rv1590 (which we named "biotin synthase auxiliary protein" or BsaP), for activity. We found homologs of bsaP associated with bioB in many actinobacterial genomes, and confirmed that BioB of Mycobacterium smegmatis also requires BsaP. Structural comparisons of BsaP-associated biotin synthases with BsaP-independent biotin synthases suggest that the need for BsaP is determined by the [2Fe-2S] cluster that inserts sulfur into dethiobiotin. Our findings open new opportunities to seek BioB inhibitors to treat infections with M. tuberculosis and other pathogens.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Global Tuberculosis Report 2021. Licence: CC BY-NC-SA 3.0 IGO (World Health Organization, 2021).
MeSH terms
Substances
Grants and funding
- AI143784/Division of Intramural Research, National Institute of Allergy and Infectious Diseases (Division of Intramural Research of the NIAID)
- AI143575/Division of Intramural Research, National Institute of Allergy and Infectious Diseases (Division of Intramural Research of the NIAID)
- P30 CA008748/CA/NCI NIH HHS/United States
- P01 AI143575/AI/NIAID NIH HHS/United States
- R01 AI143784/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
