

Recruitment of plasma cells from IL-21-dependent and IL-21-independent immune reactions to the bone marrow
- PMID: 38755157
- PMCID: PMC11099182
- DOI: 10.1038/s41467-024-48570-0
Recruitment of plasma cells from IL-21-dependent and IL-21-independent immune reactions to the bone marrow
Abstract
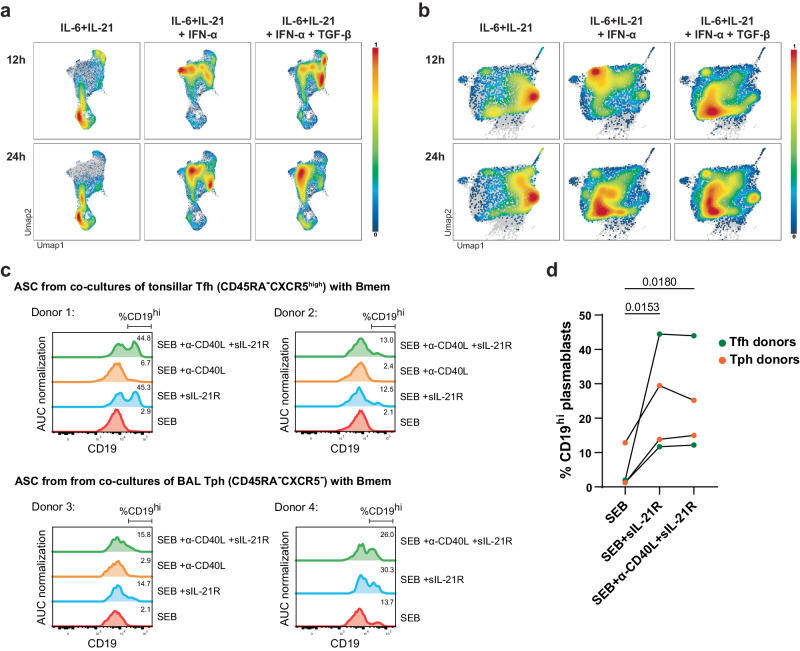
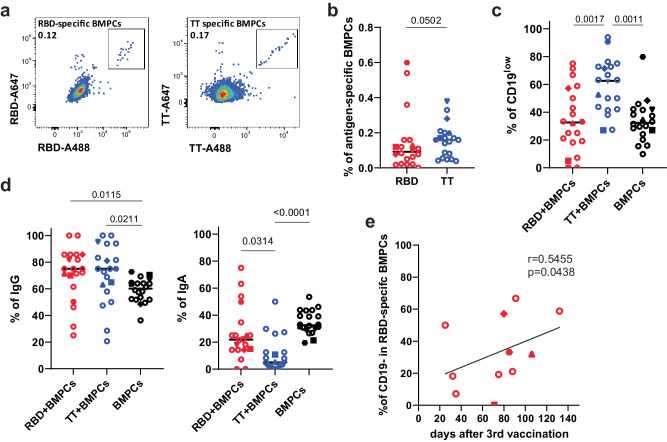
Bone marrow plasma cells (BMPC) are the correlate of humoral immunity, consistently releasing antibodies into the bloodstream. It remains unclear if BMPC reflect different activation environments or maturation of their precursors. Here we define human BMPC heterogeneity and track the recruitment of antibody-secreting cells (ASC) from SARS-CoV-2 vaccine immune reactions to the bone marrow (BM). Trajectories based on single-cell transcriptomes and repertoires of peripheral and BM ASC reveal sequential colonisation of BMPC compartments. In activated B cells, IL-21 suppresses CD19 expression, indicating that CD19low-BMPC are derived from follicular, while CD19high-BMPC originate from extrafollicular immune reactions. In primary immune reactions, both CD19low- and CD19high-BMPC compartments are populated. In secondary immune reactions, most BMPC are recruited to CD19high-BMPC compartments, reflecting their origin from extrafollicular reactivations of memory B cells. A pattern also observable in vaccinated-convalescent individuals and upon diphtheria/tetanus/pertussis recall-vaccination. Thus, BMPC diversity reflects the evolution of a given humoral immune response.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Radbruch A, et al. Competence and competition: the challenge of becoming a long-lived plasma cell. Nat. Rev. Immunol. 2006;6:741–750. - PubMed
-
- Manz RA, Thiel A, Radbruch A. Lifetime of plasma cells in the bone marrow. Nature. 1997;388:133–134. - PubMed
-
- Slifka MK, Antia R, Whitmire JK, Ahmed R. Humoral immunity due to long-lived plasma cells. Immunity. 1998;8:363–372. - PubMed
-
- Turner JS, et al. SARS-CoV-2 infection induces long-lived bone marrow plasma cells in humans. Nature. 2021;595:421–425. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
