

Hypoxia coordinates the spatial landscape of myeloid cells within glioblastoma to affect survival
- PMID: 38758780
- PMCID: PMC11100569
- DOI: 10.1126/sciadv.adj3301
Hypoxia coordinates the spatial landscape of myeloid cells within glioblastoma to affect survival
Abstract
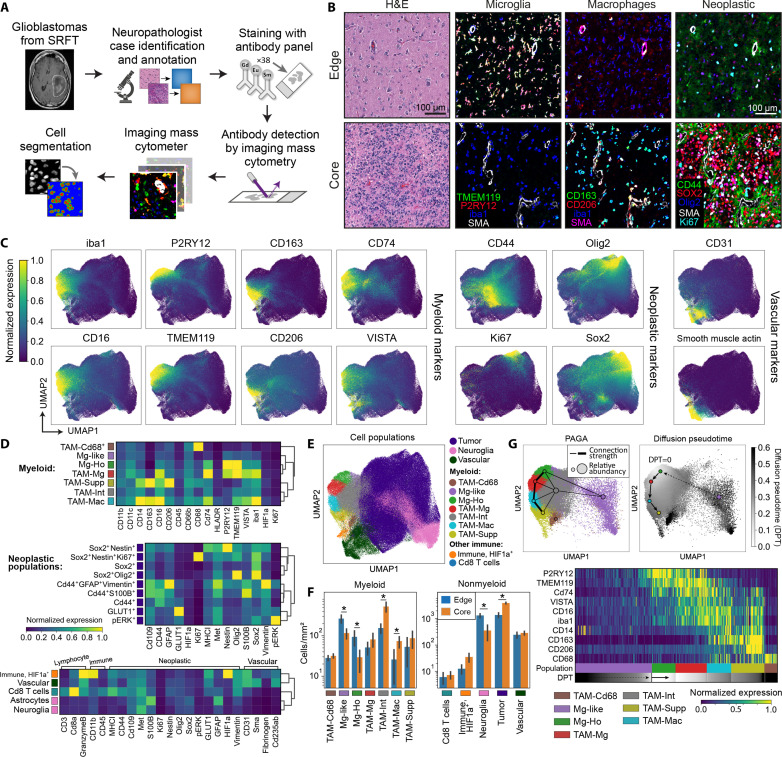
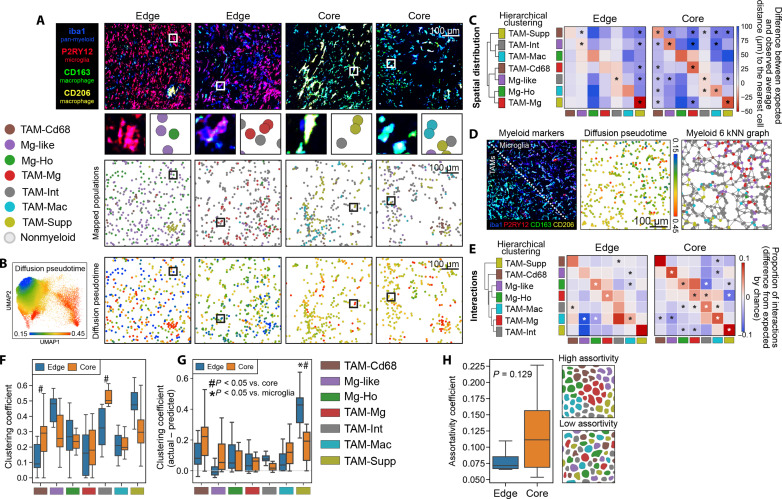
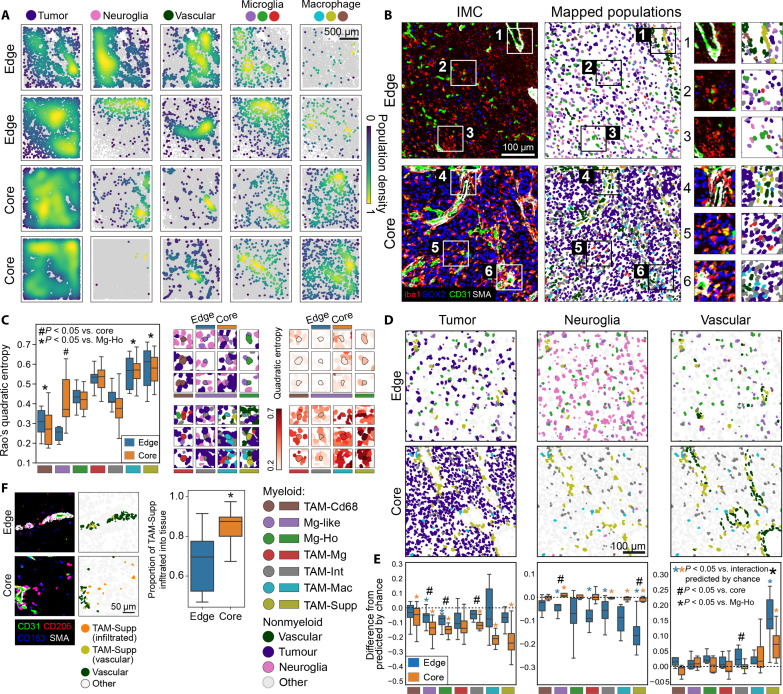
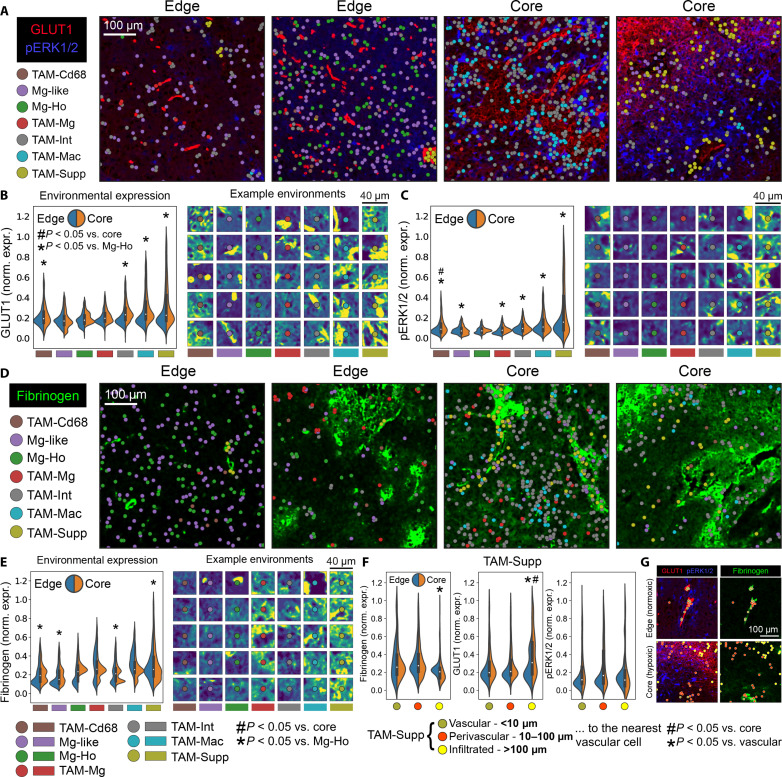
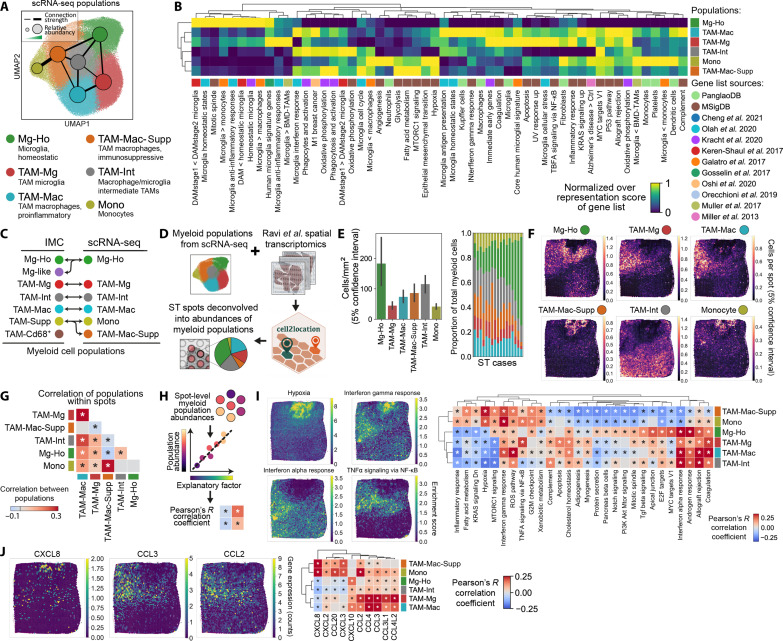
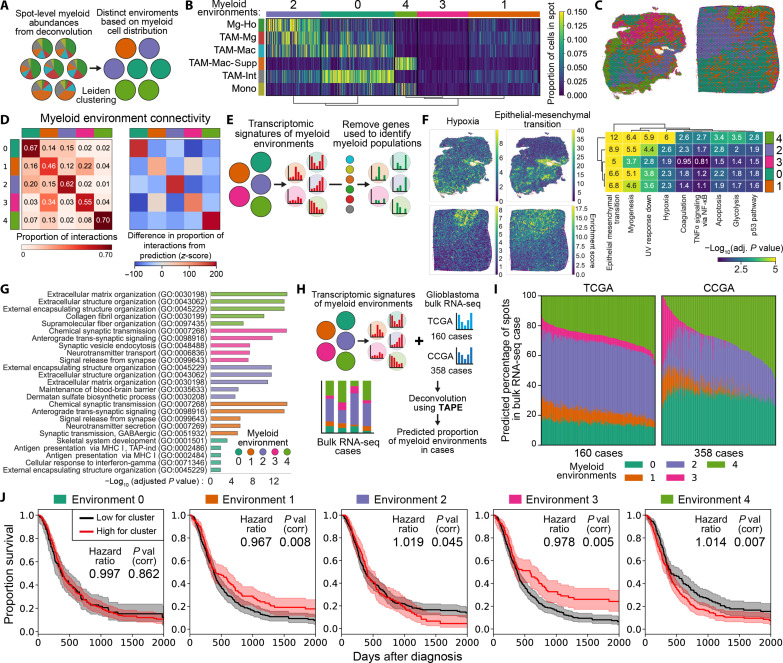
Myeloid cells are highly prevalent in glioblastoma (GBM), existing in a spectrum of phenotypic and activation states. We now have limited knowledge of the tumor microenvironment (TME) determinants that influence the localization and the functions of the diverse myeloid cell populations in GBM. Here, we have utilized orthogonal imaging mass cytometry with single-cell and spatial transcriptomic approaches to identify and map the various myeloid populations in the human GBM tumor microenvironment (TME). Our results show that different myeloid populations have distinct and reproducible compartmentalization patterns in the GBM TME that is driven by tissue hypoxia, regional chemokine signaling, and varied homotypic and heterotypic cellular interactions. We subsequently identified specific tumor subregions in GBM, based on composition of identified myeloid cell populations, that were linked to patient survival. Our results provide insight into the spatial organization of myeloid cell subpopulations in GBM, and how this is predictive of clinical outcome.
Figures
References
-
- Chuntova P., Chow F., Watchmaker P. B., Galvez M., Heimberger A. B., Newell E. W., Diaz A., DePinho R. A., Li M. O., Wherry E. J., Mitchell D., Terabe M., Wainwright D. A., Berzofsky J. A., Herold-Mende C., Heath J. R., Lim M., Margolin K. A., Chiocca E. A., Kasahara N., Ellingson B. M., Brown C. E., Chen Y., Fecci P. E., Reardon D. A., Dunn G. P., Liau L. M., Costello J. F., Wick W., Cloughesy T., Timmer W. C., Wen P. Y., Prins R. M., Platten M., Okada H., Unique challenges for glioblastoma immunotherapy—discussions across neuro-oncology and non-neuro-oncology experts in cancer immunology. Meeting Report from the 2019 SNO Immuno-Oncology Think Tank. Neuro Oncol. 23, 356–375 (2021). - PMC - PubMed
-
- Larkin C. J., Arrieta V. A., Najem H., Li G., Zhang P., Miska J., Chen P., James C. D., Sonabend A. M., Heimberger A. B., Myeloid cell classification and therapeutic opportunities within the glioblastoma tumor microenvironment in the single cell-omics era. Front. Immunol. 13, 907605 (2022). - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
