

Arginine deprivation enriches lung cancer proteomes with cysteine by inducing arginine-to-cysteine substitutants
- PMID: 38759626
- PMCID: PMC11129317
- DOI: 10.1016/j.molcel.2024.04.012
Arginine deprivation enriches lung cancer proteomes with cysteine by inducing arginine-to-cysteine substitutants
Abstract
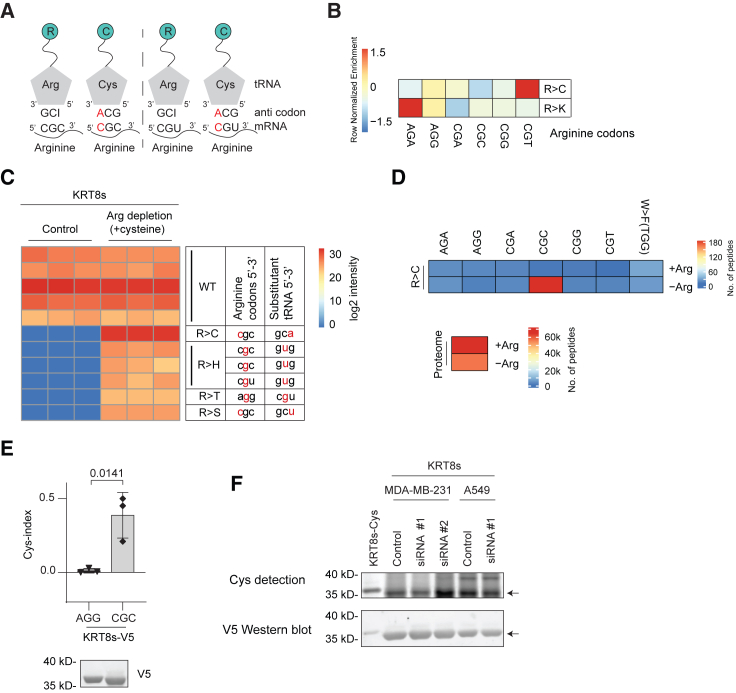
Many types of human cancers suppress the expression of argininosuccinate synthase 1 (ASS1), a rate-limiting enzyme for arginine production. Although dependency on exogenous arginine can be harnessed by arginine-deprivation therapies, the impact of ASS1 suppression on the quality of the tumor proteome is unknown. We therefore interrogated proteomes of cancer patients for arginine codon reassignments (substitutants) and surprisingly identified a strong enrichment for cysteine (R>C) in lung tumors specifically. Most R>C events did not coincide with genetically encoded R>C mutations but were likely products of tRNA misalignments. The expression of R>C substitutants was highly associated with oncogenic kelch-like epichlorohydrin (ECH)-associated protein 1 (KEAP1)-pathway mutations and suppressed by intact-KEAP1 in KEAP1-mutated cancer cells. Finally, functional interrogation indicated a key role for R>C substitutants in cell survival to cisplatin, suggesting that regulatory codon reassignments endow cancer cells with more resilience to stress. Thus, we present a mechanism for enriching lung cancer proteomes with cysteines that may affect therapeutic decisions.
Keywords: aberrant mRNA translation; amino acid shortage; arginine deprivation; chemotherapy; cysteine; ferroptosis; lung cancer; substitutants.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures







References
-
- Ganjoo S., Gupta P., Corbali H.I., Nanez S., Riad T.S., Duong L.K., Barsoumian H.B., Masrorpour F., Jiang H., Welsh J.W., Cortez M.A. The role of tumor metabolism in modulating T-Cell activity and in optimizing immunotherapy. Front. Immunol. 2023;14 doi: 10.3389/fimmu.2023.1172931. - DOI - PMC - PubMed
-
- Nakaya M., Xiao Y., Zhou X., Chang J.H., Chang M., Cheng X., Blonska M., Lin X., Sun S.C. Inflammatory T cell responses rely on amino acid transporter ASCT2 facilitation of glutamine uptake and mTORC1 kinase activation. Immunity. 2014;40:692–705. doi: 10.1016/j.immuni.2014.04.007. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
