

Methylmalonic acidemia triggers lysosomal-autophagy dysfunctions
- PMID: 38760822
- PMCID: PMC11102240
- DOI: 10.1186/s13578-024-01245-1
Methylmalonic acidemia triggers lysosomal-autophagy dysfunctions
Abstract
Background: Methylmalonic acidemia (MMA) is a rare inborn error of propionate metabolism caused by deficiency of the mitochondrial methylmalonyl-CoA mutase (MUT) enzyme. As matter of fact, MMA patients manifest impairment of the primary metabolic network with profound damages that involve several cell components, many of which have not been discovered yet. We employed cellular models and patients-derived fibroblasts to refine and uncover new pathologic mechanisms connected with MUT deficiency through the combination of multi-proteomics and bioinformatics approaches.
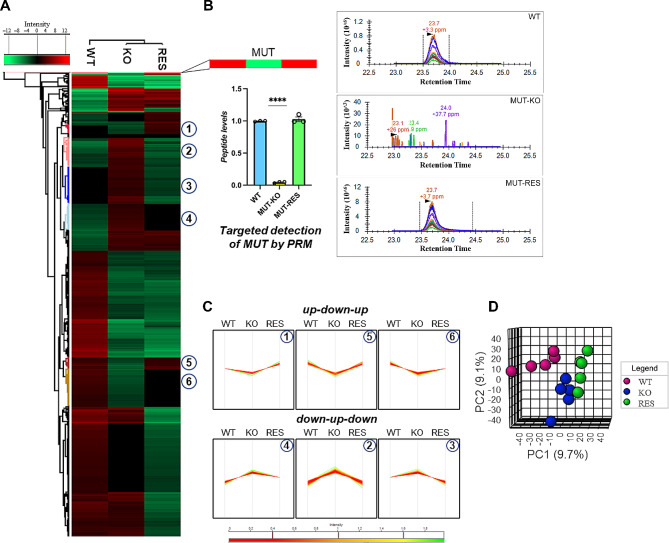
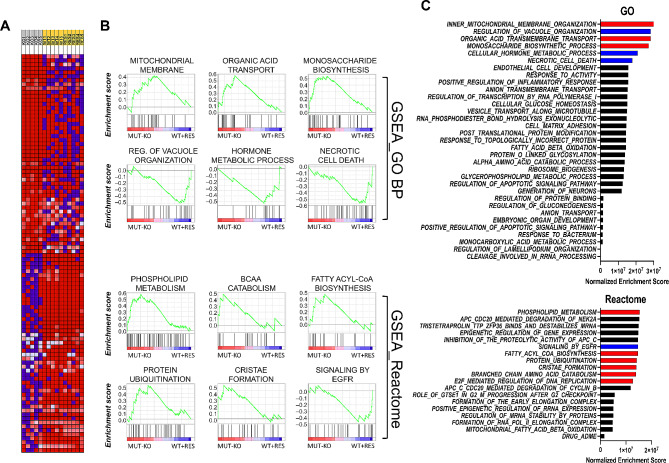
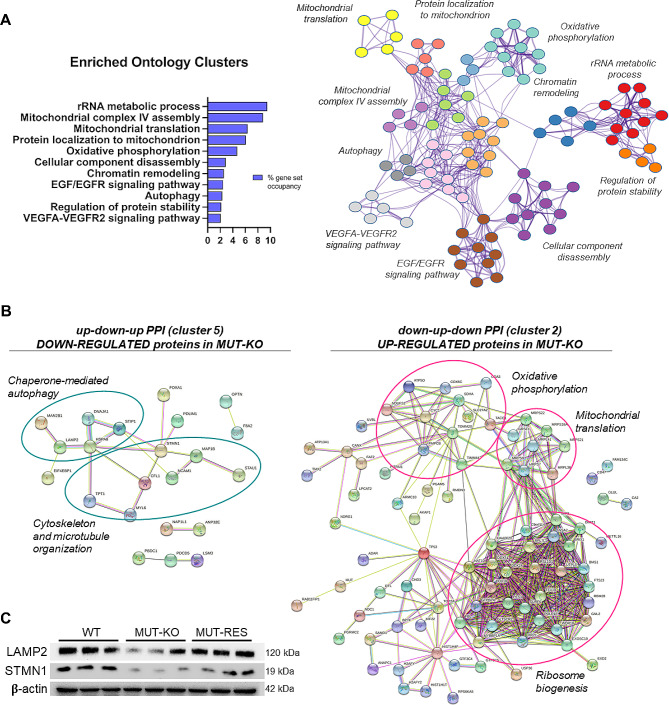
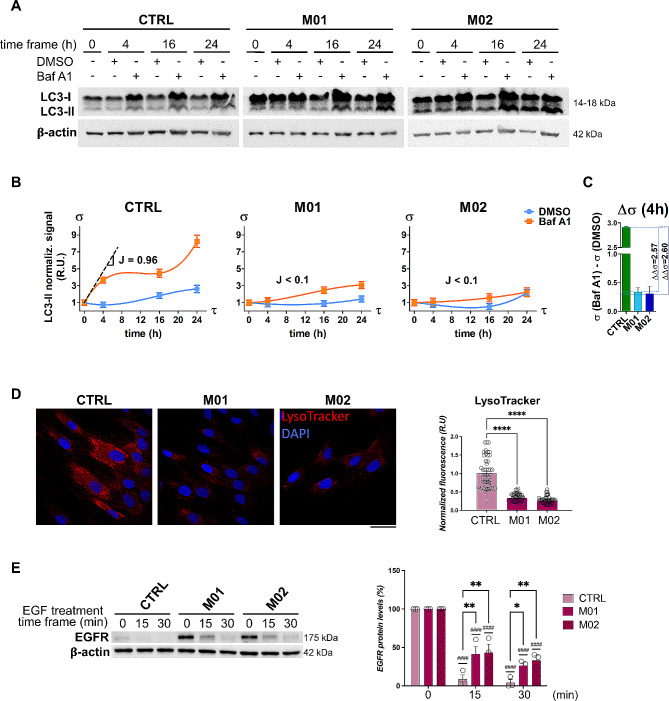
Results: Our data show that MUT deficiency is connected with profound proteome dysregulations, revealing molecular actors involved in lysosome and autophagy functioning. To elucidate the effects of defective MUT on lysosomal and autophagy regulation, we analyzed the morphology and functionality of MMA-lysosomes that showed deep alterations, thus corroborating omics data. Lysosomes of MMA cells present as enlarged vacuoles with low degradative capabilities. Notwithstanding, treatment with an anti-propionigenic drug is capable of totally rescuing lysosomal morphology and functional activity in MUT-deficient cells. These results indicate a strict connection between MUT deficiency and lysosomal-autophagy dysfunction, providing promising therapeutic perspectives for MMA.
Conclusions: Defective homeostatic mechanisms in the regulation of autophagy and lysosome functions have been demonstrated in MUT-deficient cells. Our data prove that MMA triggers such dysfunctions impacting on autophagosome-lysosome fusion and lysosomal activity.
Keywords: Autophagy; Lysosomes; MMA therapy; Metabolic disease; Methylmalonic acidemia; Multi-omics; Multi-proteomics.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no potential competing interests.
Figures


References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
