

TBC9, an essential TBC-domain protein, regulates early vesicular transport and IMC formation in Toxoplasma gondii
- PMID: 38762629
- PMCID: PMC11102469
- DOI: 10.1038/s42003-024-06310-6
TBC9, an essential TBC-domain protein, regulates early vesicular transport and IMC formation in Toxoplasma gondii
Abstract
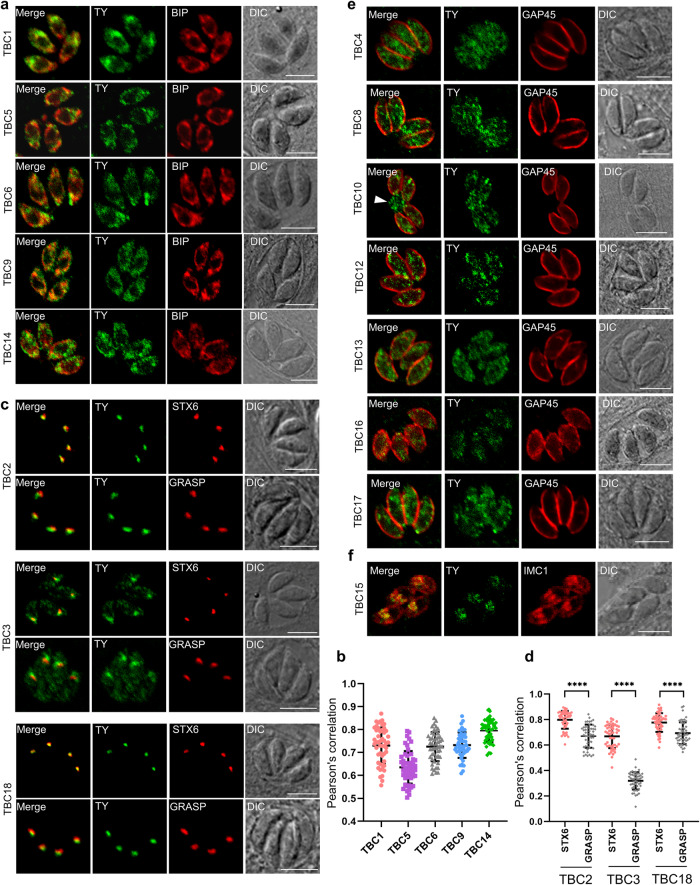
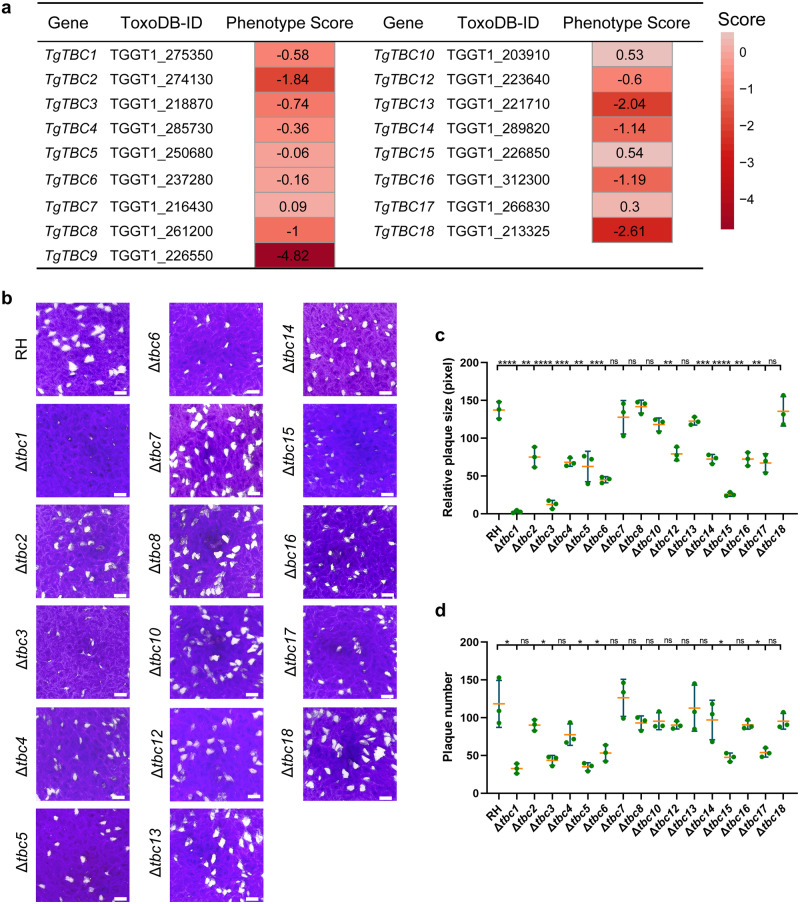
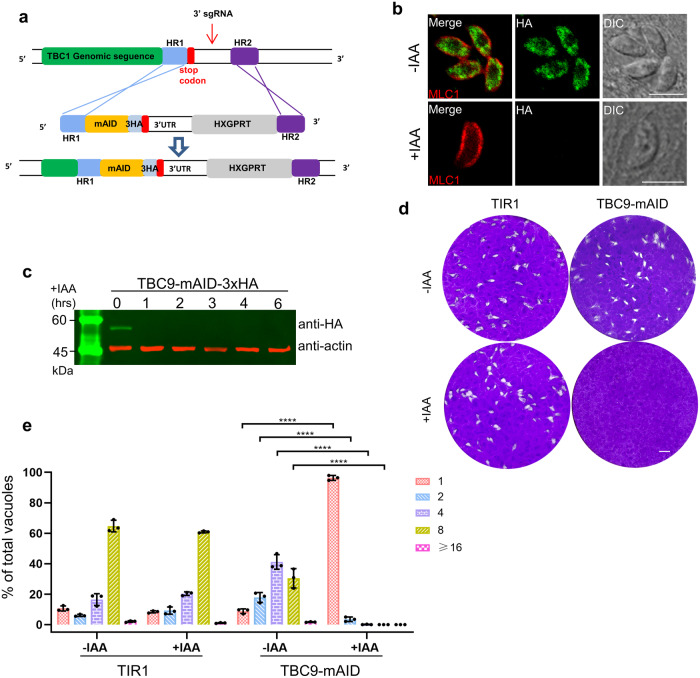
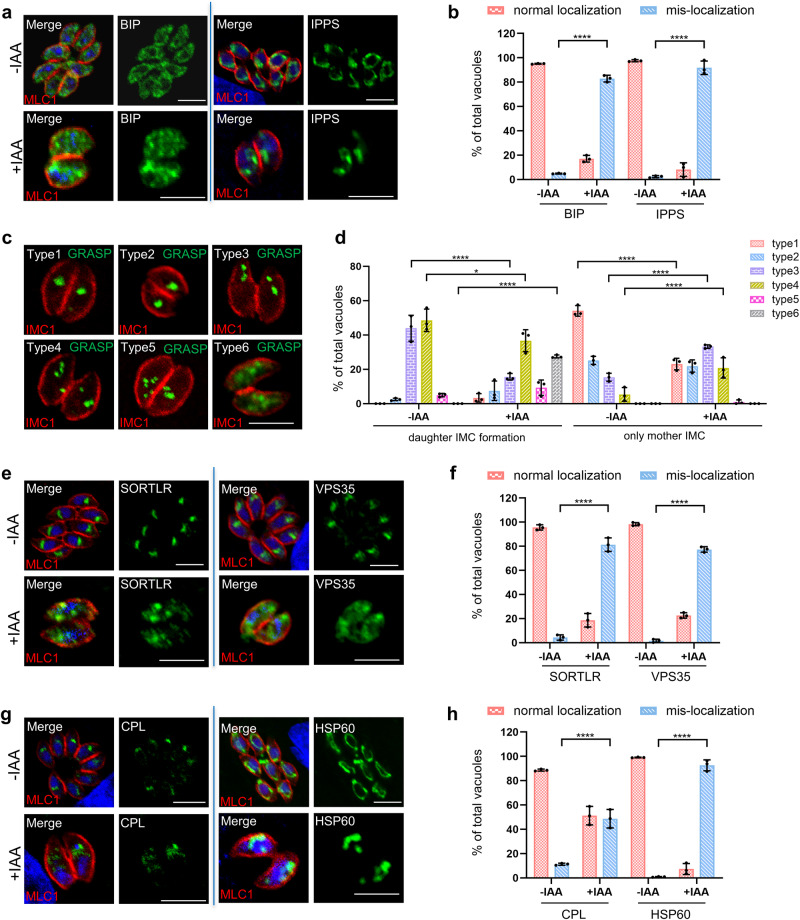
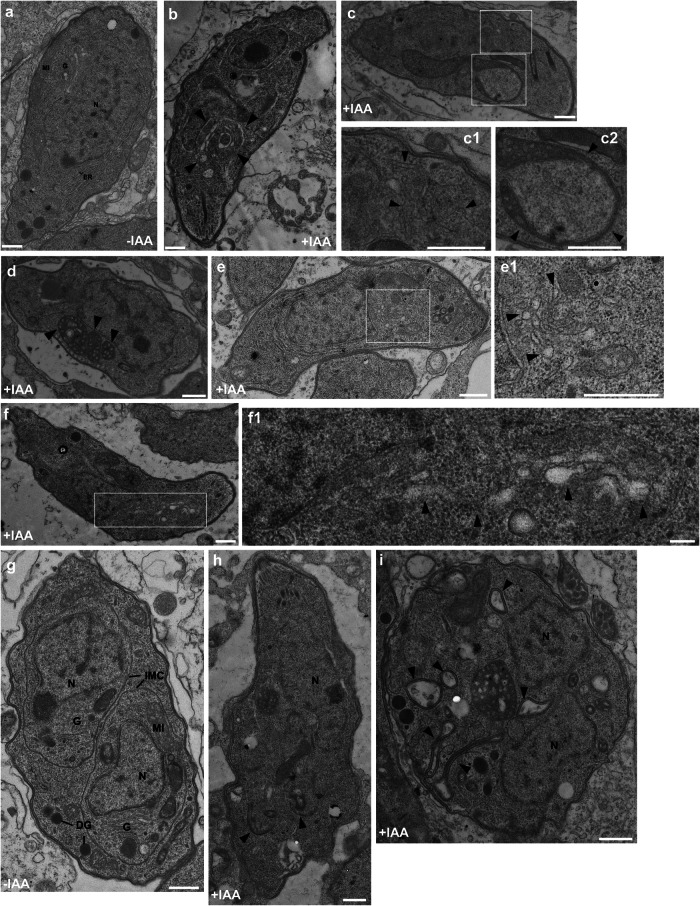
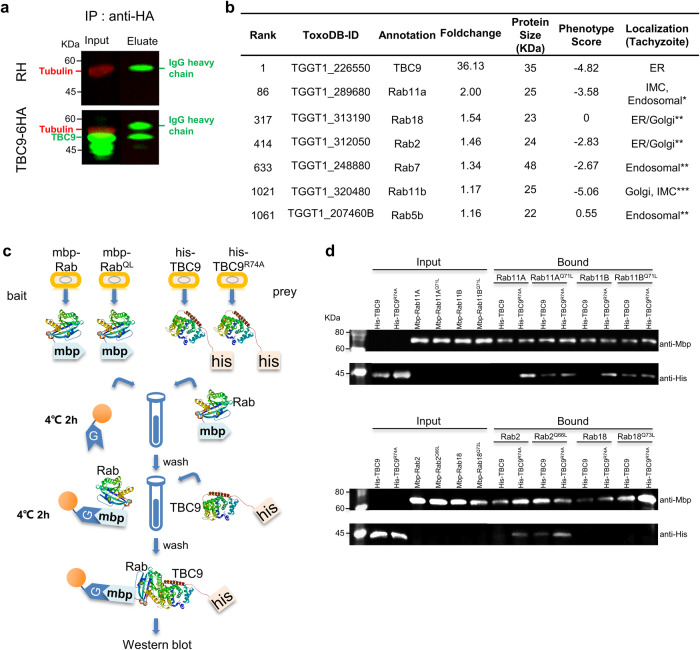
Apicomplexan parasites harbor a complex endomembrane system as well as unique secretory organelles. These complex cellular structures require an elaborate vesicle trafficking system, which includes Rab GTPases and their regulators, to assure the biogenesis and secretory of the organelles. Here we exploit the model apicomplexan organism Toxoplasma gondii that encodes a family of Rab GTPase Activating Proteins, TBC (Tre-2/Bub2/Cdc16) domain-containing proteins. Functional profiling of these proteins in tachyzoites reveals that TBC9 is the only essential regulator, which is localized to the endoplasmic reticulum (ER) in T. gondii strains. Detailed analyses demonstrate that TBC9 is required for normal distribution of proteins targeting to the ER, and the Golgi apparatus in the parasite, as well as for the normal formation of daughter inner membrane complexes (IMCs). Pull-down assays show a strong protein interaction between TBC9 and specific Rab GTPases (Rab11A, Rab11B, and Rab2), supporting the role of TBC9 in daughter IMC formation and early vesicular transport. Thus, this study identifies the only essential TBC domain-containing protein TBC9 that regulates early vesicular transport and IMC formation in T. gondii and potentially in closely related protists.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Systematic characterization of all Toxoplasma gondii TBC domain-containing proteins identifies an essential regulator of Rab2 in the secretory pathway.PLoS Biol. 2024 May 7;22(5):e3002634. doi: 10.1371/journal.pbio.3002634. eCollection 2024 May. PLoS Biol. 2024. PMID: 38713739 Free PMC article.
-
The Dissection of SNAREs Reveals Key Factors for Vesicular Trafficking to the Endosome-like Compartment and Apicoplast via the Secretory System in Toxoplasma gondii.mBio. 2021 Aug 31;12(4):e0138021. doi: 10.1128/mBio.01380-21. Epub 2021 Aug 3. mBio. 2021. PMID: 34340555 Free PMC article.
-
Biogenesis of the inner membrane complex is dependent on vesicular transport by the alveolate specific GTPase Rab11B.PLoS Pathog. 2010 Jul 29;6(7):e1001029. doi: 10.1371/journal.ppat.1001029. PLoS Pathog. 2010. PMID: 20686666 Free PMC article.
-
Rab proteins of the endoplasmic reticulum: functions and interactors.Biochem Soc Trans. 2012 Dec 1;40(6):1426-32. doi: 10.1042/BST20120158. Biochem Soc Trans. 2012. PMID: 23176493 Review.
-
Protein trafficking inside Toxoplasma gondii.Traffic. 2008 May;9(5):636-46. doi: 10.1111/j.1600-0854.2008.00713.x. Epub 2008 Mar 4. Traffic. 2008. PMID: 18331382 Review.
Cited by
-
CRISPR screens identify genes essential for in vivo virulence among proteins of hyperLOPIT-unassigned subcellular localization in Toxoplasma.mBio. 2024 Sep 11;15(9):e0172824. doi: 10.1128/mbio.01728-24. Epub 2024 Jul 31. mBio. 2024. PMID: 39082802 Free PMC article.
-
Functional dissection of prenyltransferases reveals roles in endocytosis and secretory vacuolar sorting in type 2-ME49 strain of Toxoplasma gondii.Virulence. 2024 Dec;15(1):2432681. doi: 10.1080/21505594.2024.2432681. Epub 2024 Nov 26. Virulence. 2024. PMID: 39569525 Free PMC article.
-
Role of Toxoplasma gondii p24δ in Regulating the Transition from Tachyzoite to Bradyzoite Development.Int J Mol Sci. 2025 Apr 3;26(7):3331. doi: 10.3390/ijms26073331. Int J Mol Sci. 2025. PMID: 40244171 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
