

Dorsolateral septum GLP-1R neurons regulate feeding via lateral hypothalamic projections
- PMID: 38763494
- PMCID: PMC11153235
- DOI: 10.1016/j.molmet.2024.101960
Dorsolateral septum GLP-1R neurons regulate feeding via lateral hypothalamic projections
Abstract
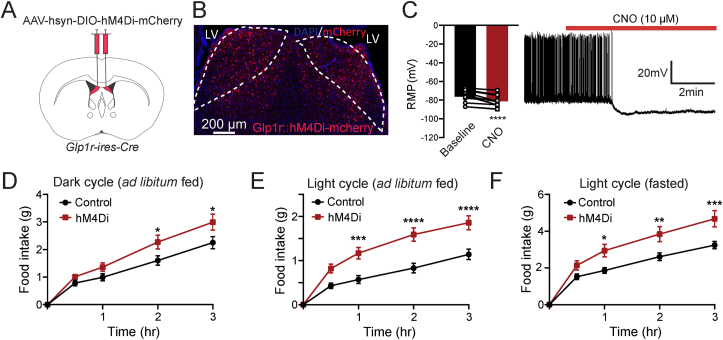
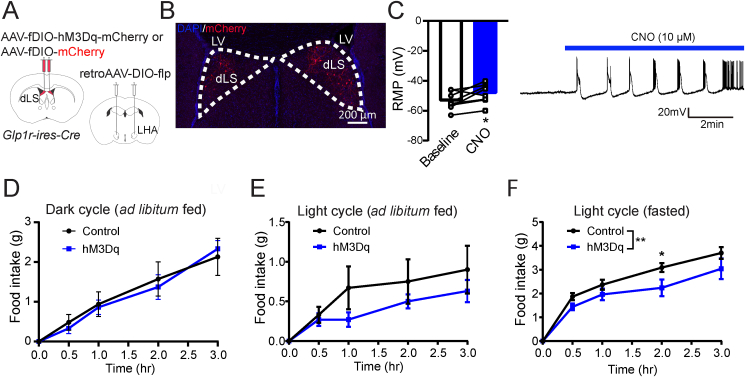
Objective: Although glucagon-like peptide 1 (GLP-1) is known to regulate feeding, the central mechanisms contributing to this function remain enigmatic. Here, we aim to test the role of neurons expressing GLP-1 receptors (GLP-1R) in the dorsolateral septum (dLS; dLSGLP-1R) that project to the lateral hypothalamic area (LHA) on food intake and determine the relationship with feeding regulation.
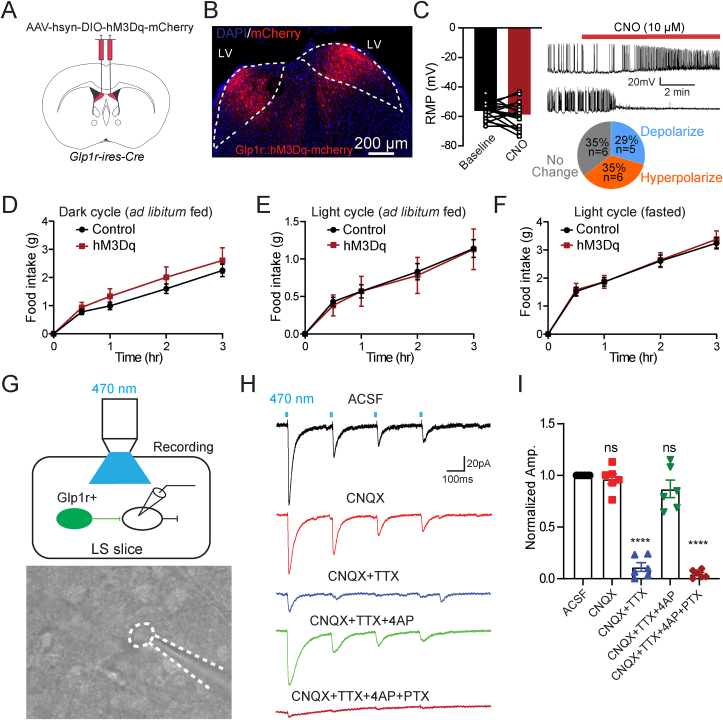
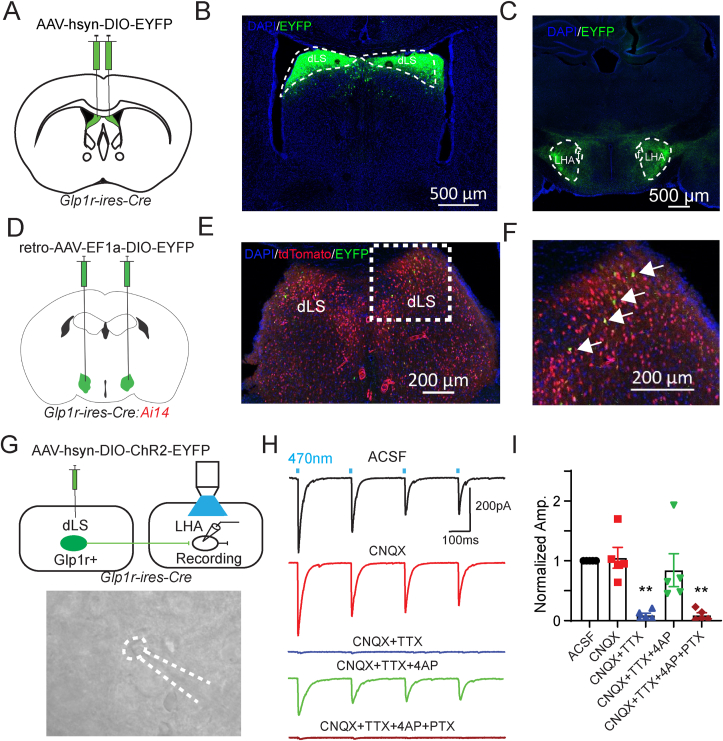
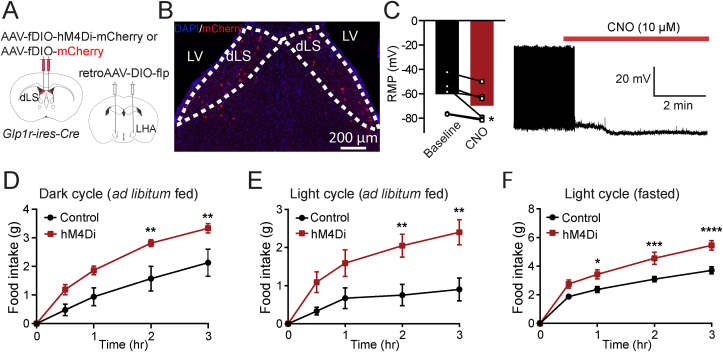
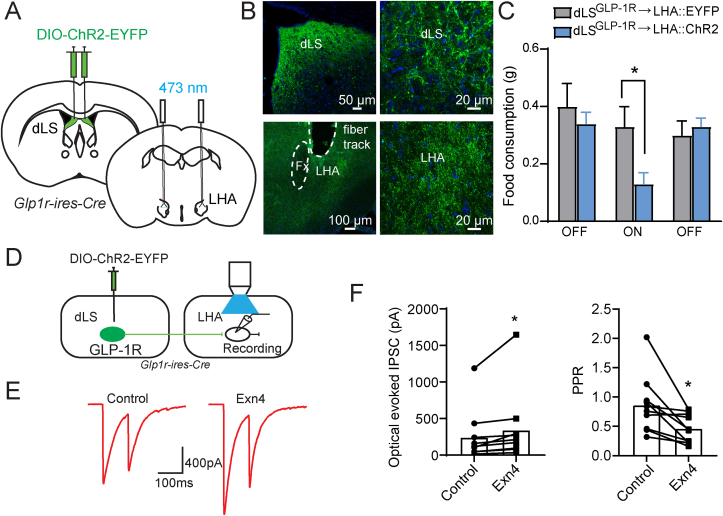
Methods: Using chemogenetic manipulations, we assessed how activation or inhibition of dLSGLP-1R neurons affected food intake in Glp1r-ires-Cre mice. Then, we used channelrhodopsin-assisted circuit mapping, chemogenetics, and electrophysiological recordings to identify and assess the role of the pathway from dLSGLP-1R →LHA projections in regulating food intake.
Results: Chemogenetic inhibition of dLSGLP-1R neurons increases food intake. LHA is a major downstream target of dLSGLP-1R neurons. The dLSGLP-1R→LHA projections are GABAergic, and chemogenetic inhibition of this pathway also promotes food intake. While chemogenetic activation of dLSGLP-1R→LHA projections modestly decreases food intake, optogenetic stimulation of the dLSGLP-1R→LHA projection terminals in the LHA rapidly suppresses feeding behavior. Finally, we demonstrate that the GLP-1R agonist, Exendin 4 enhances dLSGLP-1R →LHA GABA release.
Conclusions: Together, these results demonstrate that dLS-GLP-1R neurons and the inhibitory pathway to LHA can regulate feeding behavior, which might serve as a potential therapeutic target for the treatment of eating disorders or obesity.
Keywords: Feeding; GLP-1; Hypothalamus; Obesity; Septum.
Copyright © 2024 The Authors. Published by Elsevier GmbH.. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Update of
-
Dorsolateral septum GLP-1R neurons regulate feeding via lateral hypothalamic projections.bioRxiv [Preprint]. 2024 Mar 27:2024.03.26.586855. doi: 10.1101/2024.03.26.586855. bioRxiv. 2024. Update in: Mol Metab. 2024 Jul;85:101960. doi: 10.1016/j.molmet.2024.101960. PMID: 38585874 Free PMC article. Updated. Preprint.
References
-
- Drucker D.J. Mechanisms of action and therapeutic application of glucagon-like peptide-1. Cell Metabol. 2018;27(4):740–756. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
