

Post-transcriptional (re)programming of B lymphocyte development: From bench to bedside?
- PMID: 38763703
- PMCID: PMC12320199
- DOI: 10.1016/bs.ai.2024.03.003
Post-transcriptional (re)programming of B lymphocyte development: From bench to bedside?
Abstract
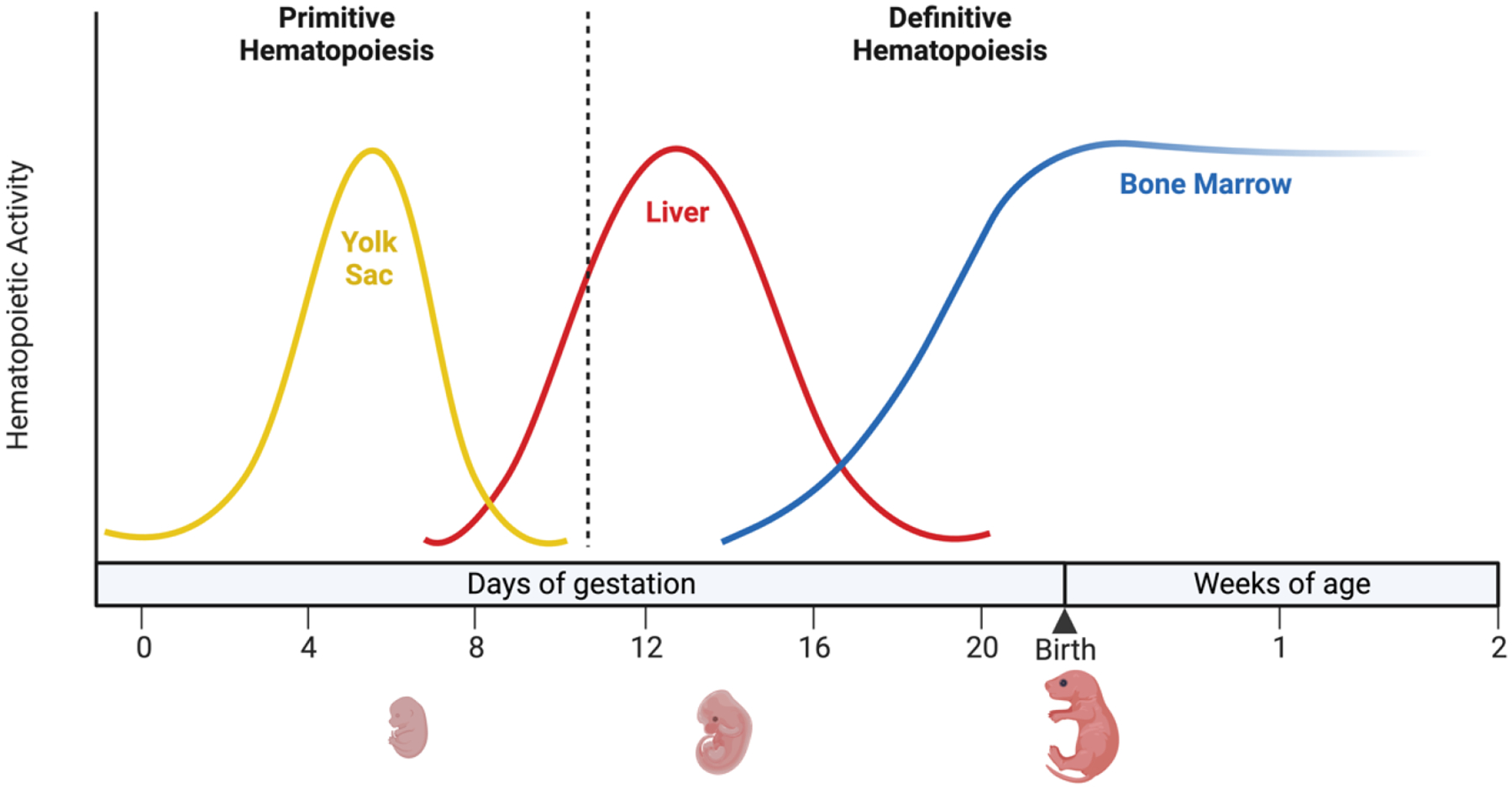
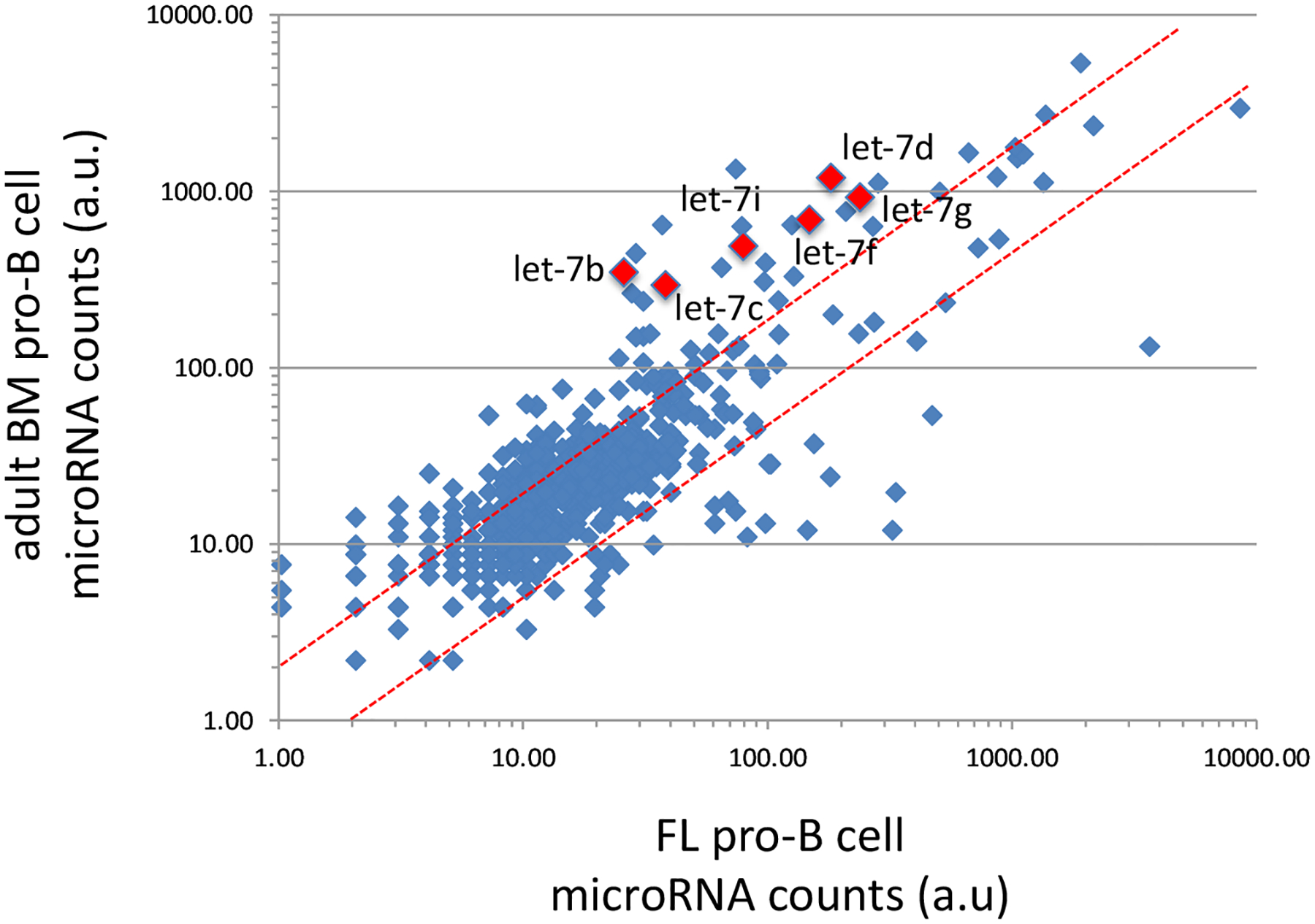
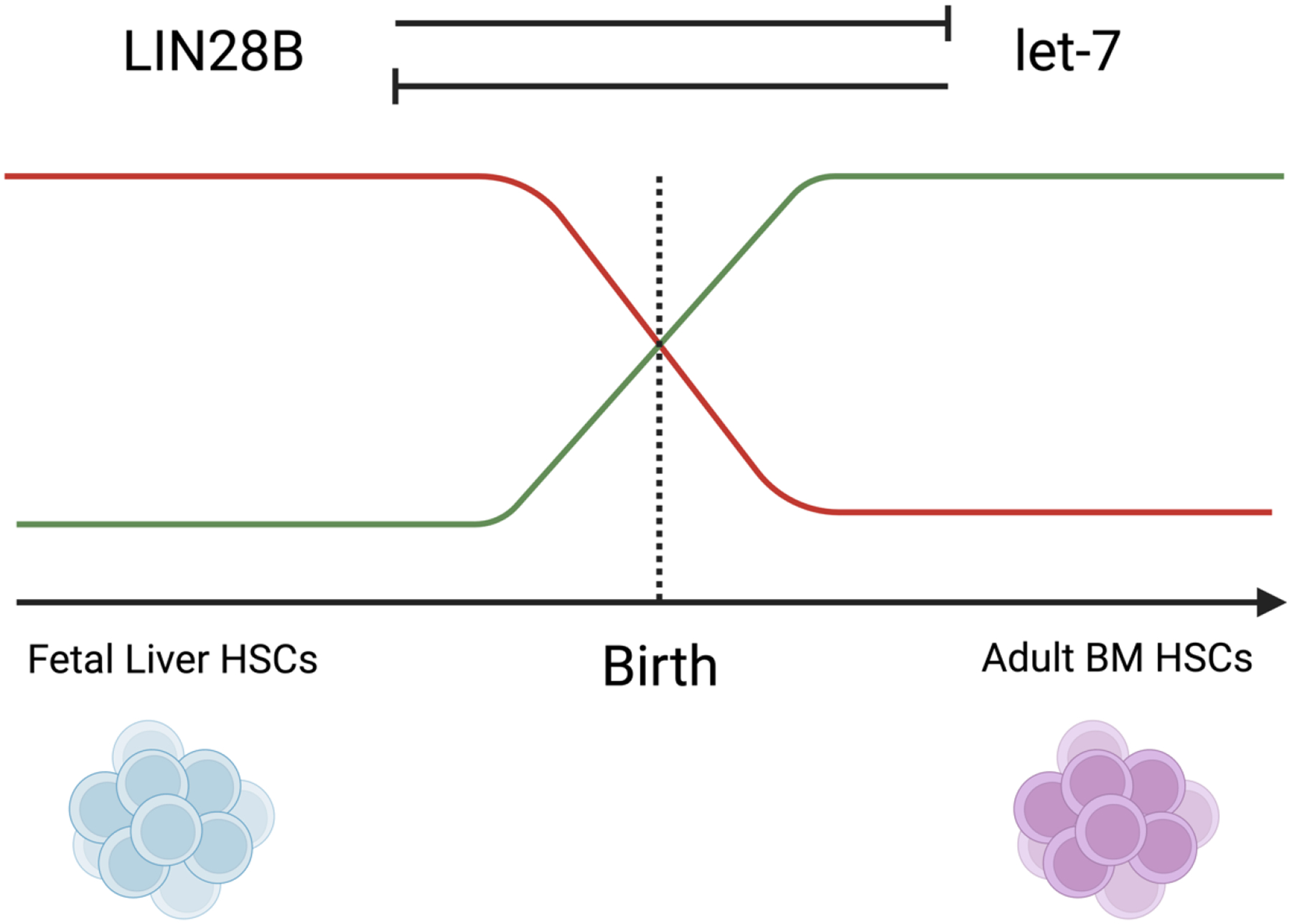
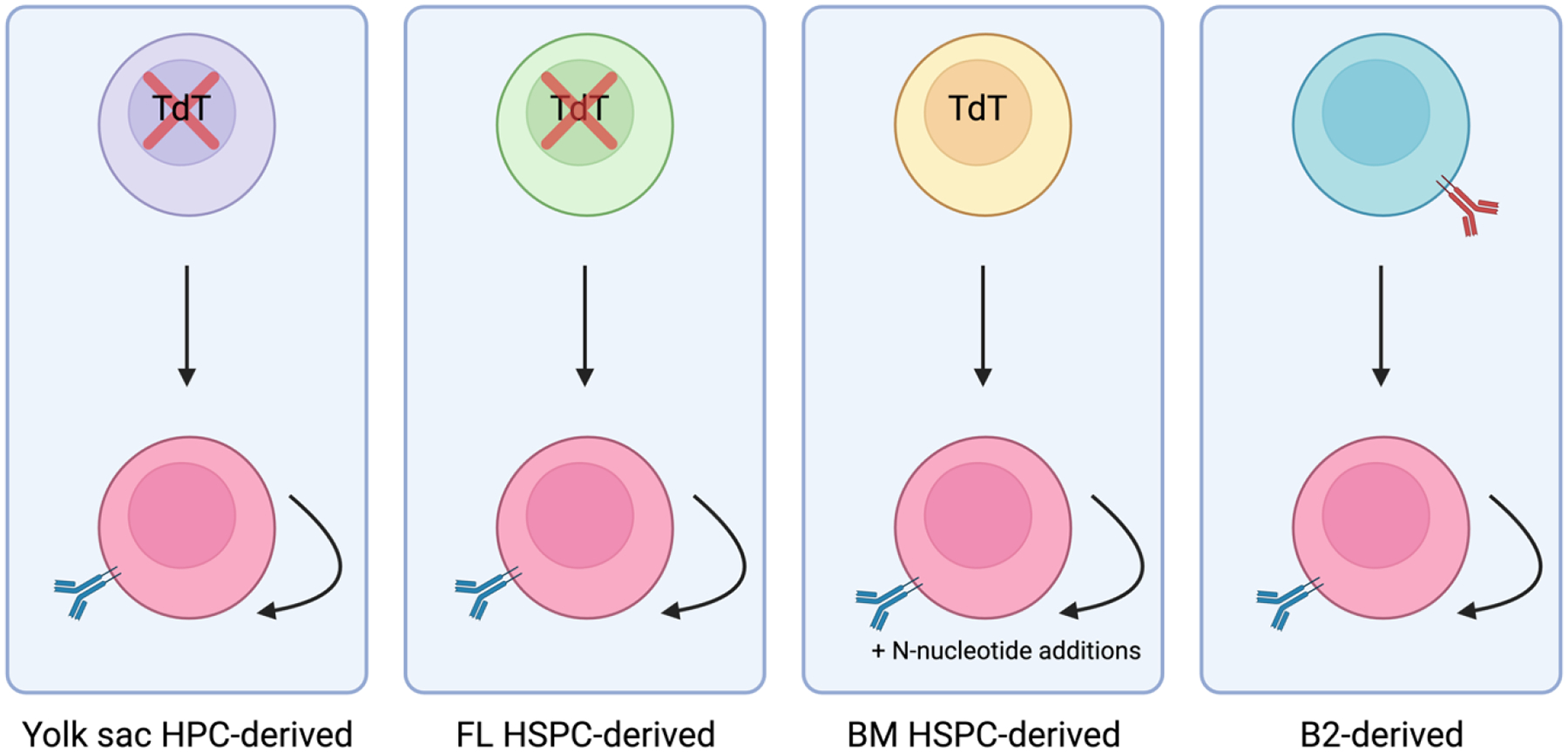
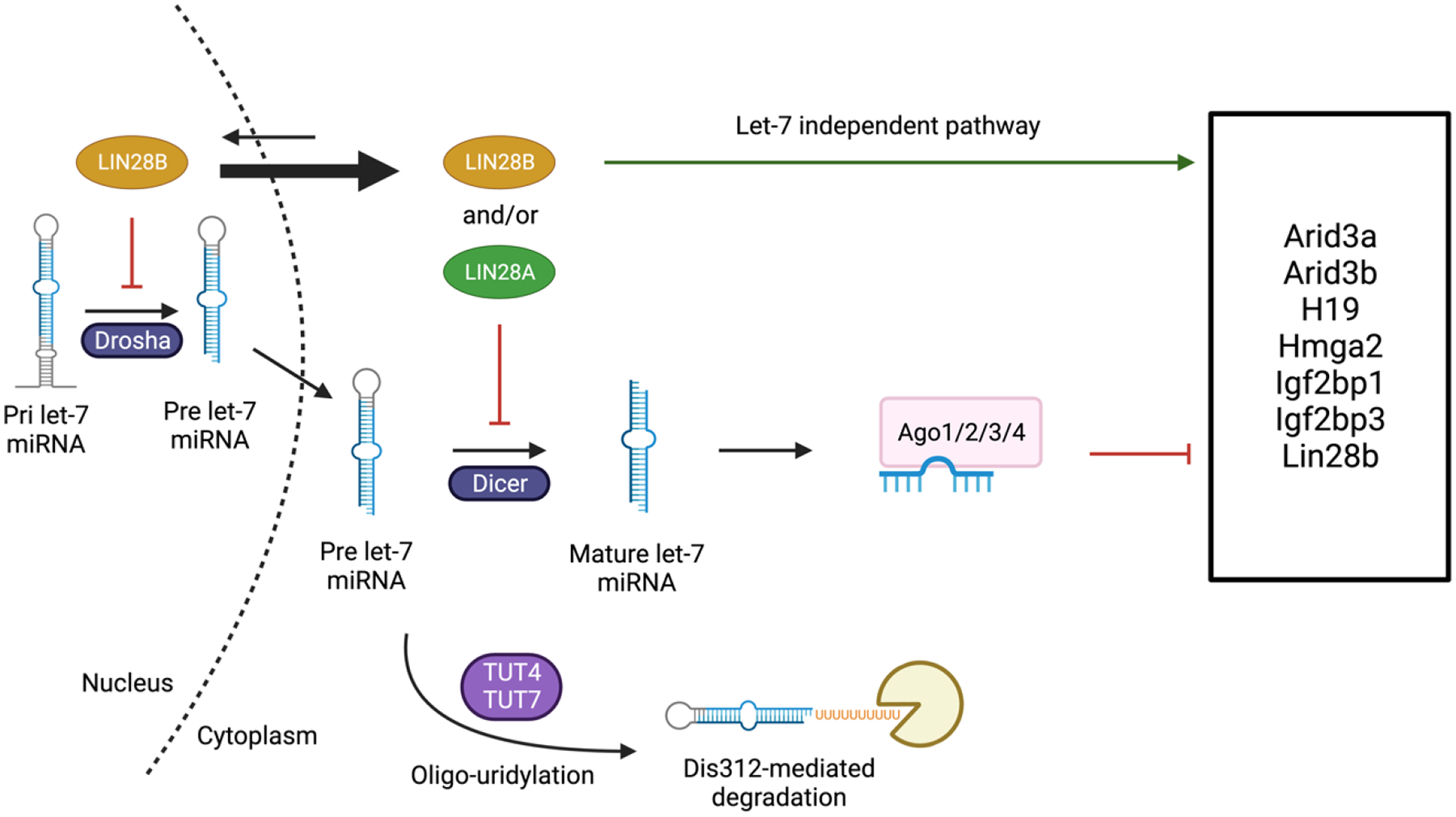
Hematopoiesis, a process which generates blood and immune cells, changes significantly during mammalian development. Definitive hematopoiesis is marked by the emergence of long-term hematopoietic stem cells (HSCs). Here, we will focus on the post-transcriptional differences between fetal liver (FL) and adult bone marrow (ABM) HSCs. It remains unclear how or why exactly FL HSCs transition to ABM HSCs, but we aim to leverage their differences to revive an old idea: in utero HSC transplantation. Unexpectedly, the expression of certain RNA-binding proteins (RBPs) play an important role in HSC specification, and can be employed to convert or reprogram adult HSCs back to a fetal-like state. Among other features, FL HSCs have a broad differentiation capacity that includes the ability to regenerate both conventional B and T cells, as well as innate-like or unconventional lymphocytes such as B-1a and marginal zone B (MzB) cells. This chapter will focus on RNA binding proteins, namely LIN28B and IGF2BP3, that are expressed during fetal life and how they promote B-1a cell development. Furthermore, this chapter considers a potential clinical application of synthetic co-expression of LIN28B and IGF2BP3 in HSCs.
Keywords: B lymphocyte development; Fetal hematopoiesis; Hematopoietic stem cells; In utero transplantation; Innate-like lymphocytes; MicroRNA; Post-transcriptional regulation; RNA-binding protein.
Copyright © 2024. Published by Elsevier Inc.
Figures
Similar articles
-
Microbiota Signals Suppress B Lymphopoiesis With Aging in Mice.Front Immunol. 2021 Oct 19;12:767267. doi: 10.3389/fimmu.2021.767267. eCollection 2021. Front Immunol. 2021. PMID: 34737755 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Regulation of HSC development and function by Lin28b.Front Cell Dev Biol. 2025 Mar 12;13:1555877. doi: 10.3389/fcell.2025.1555877. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40143971 Free PMC article. Review.
-
Exploring Epigenetic Complexity in Regulation of Hematopoietic Stem Cells Niche: A Mechanistic Journey from Normal to Malignant Hematopoiesis.Adv Exp Med Biol. 2025;1483:49-67. doi: 10.1007/5584_2024_846. Adv Exp Med Biol. 2025. PMID: 39841383 Review.
References
-
- Alejo-Valle O, Weigert K, Bhayadia R, Ng M, Issa H, Beyer C, Emmrich S, Schuschel K, Ihling C, Sinz A, Zimmermann M, Wickenhauser C, Flasinski M, Regenyi E, Labuhn M, Reinhardt D, Yaspo ML, Heckl D, & Klusmann JH (2022). The megakaryocytic transcription factor ARID3A suppresses leukemia pathogenesis. Blood, 139(5), 651–665. 10.1182/blood.2021012231 - DOI - PMC - PubMed
-
- Amabile G, Welner RS, Nombela-Arrieta C, D’Alise AM, Di Ruscio A, Ebralidze AK, Kraytsberg Y, Ye M, Kocher O, Neuberg DS, Khrapko K, Silberstein LE, & Tenen DG (2013). In vivo generation of transplantable human hematopoietic cells from induced pluripotent stem cells. Blood, 121(8), 1255–1264. 10.1182/blood-2012-06-434407 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
