

This is a preprint.
Glycosaminoglycan-mediated lipoprotein uptake protects cancer cells from ferroptosis
- PMID: 38765991
- PMCID: PMC11101130
- DOI: 10.1101/2024.05.13.593939
Glycosaminoglycan-mediated lipoprotein uptake protects cancer cells from ferroptosis
Update in
-
Glycosaminoglycan-driven lipoprotein uptake protects tumours from ferroptosis.Nature. 2025 Aug;644(8077):799-808. doi: 10.1038/s41586-025-09162-0. Epub 2025 Jun 11. Nature. 2025. PMID: 40500442
Abstract
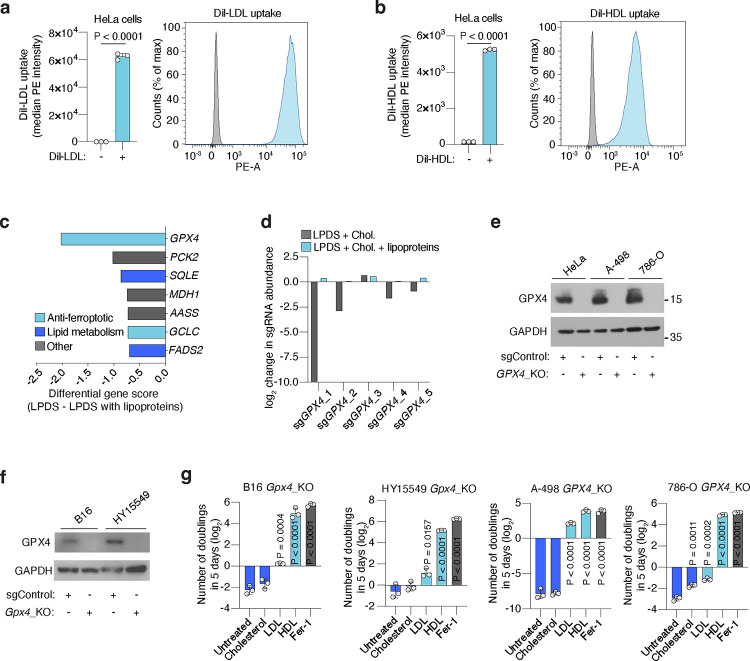
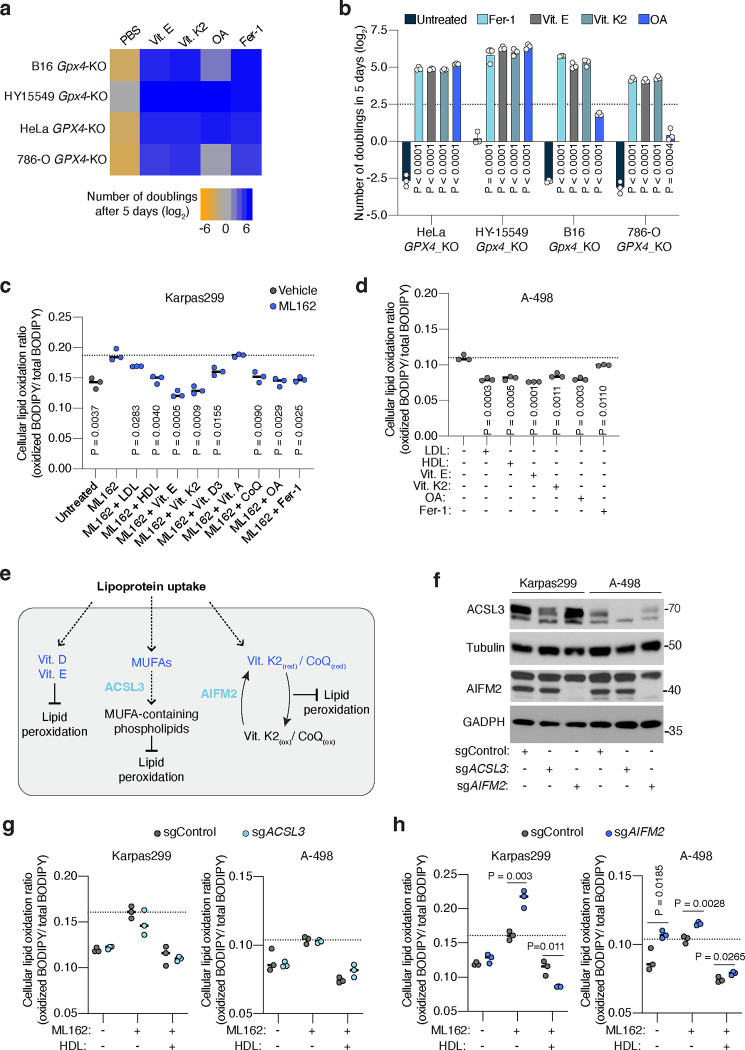
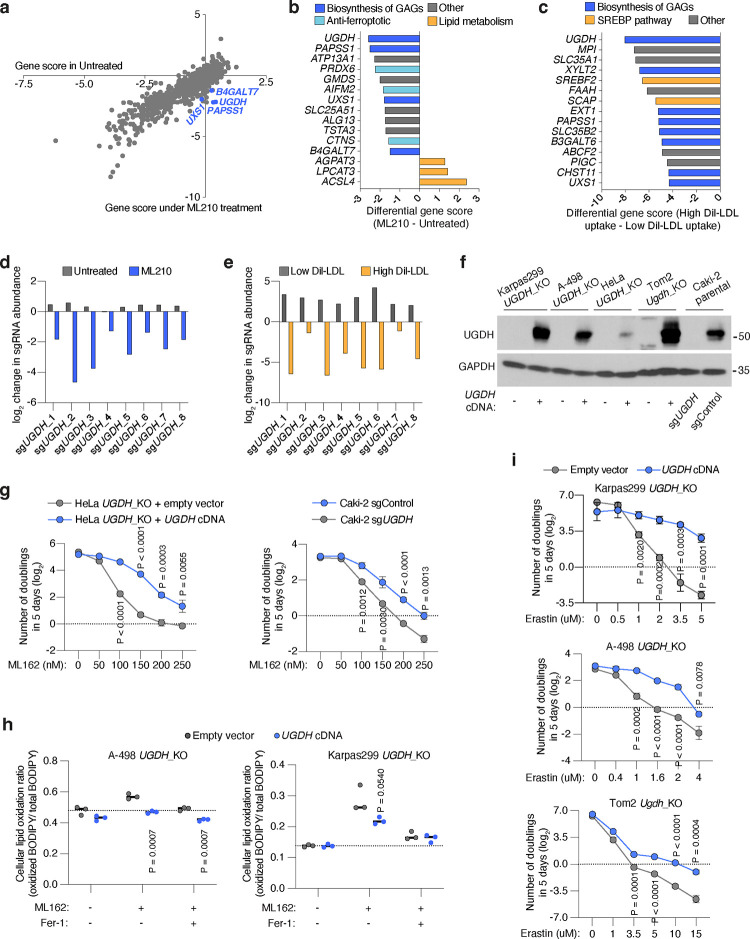
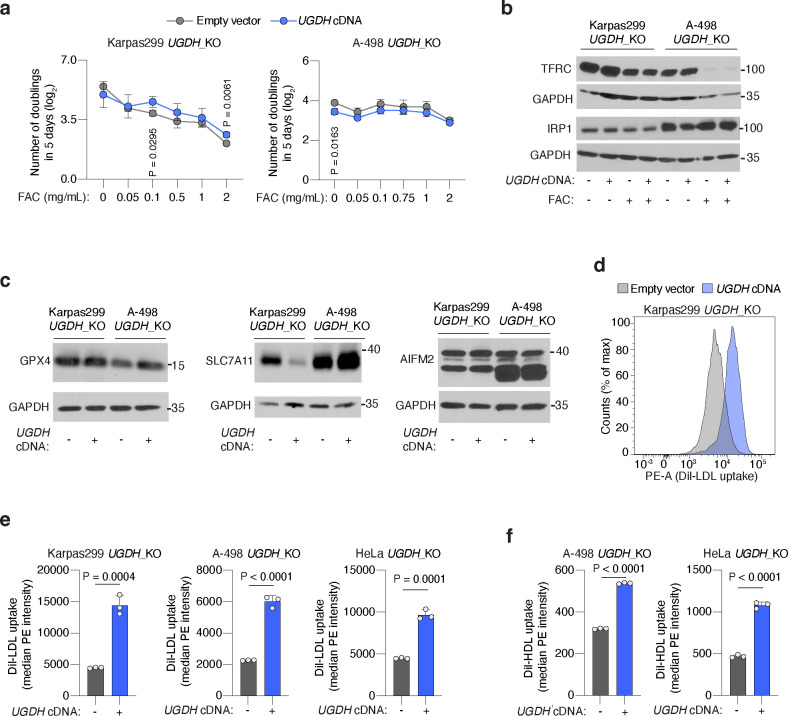
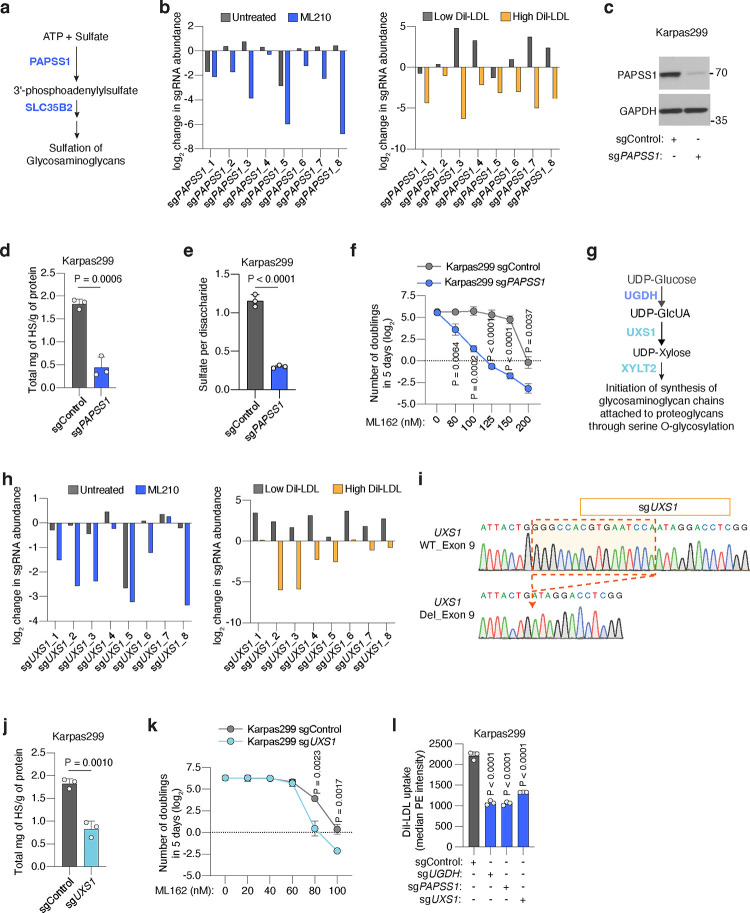
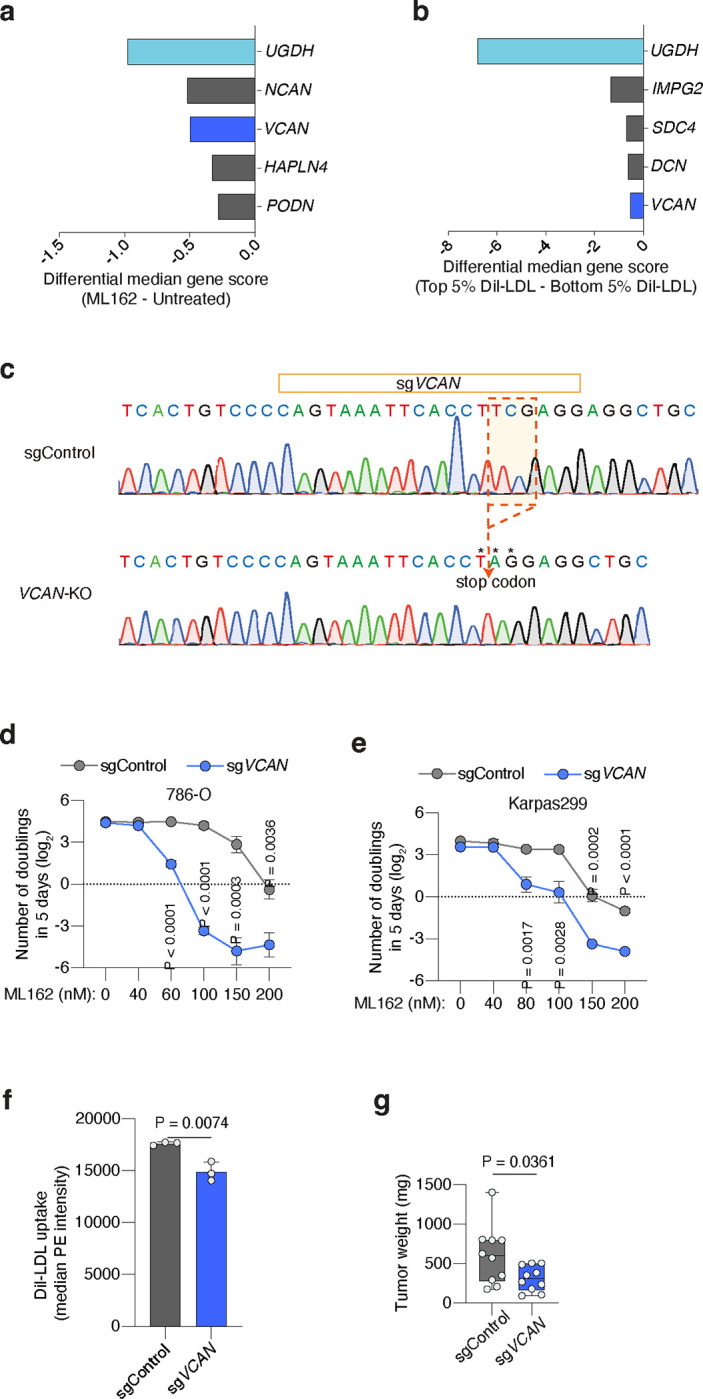
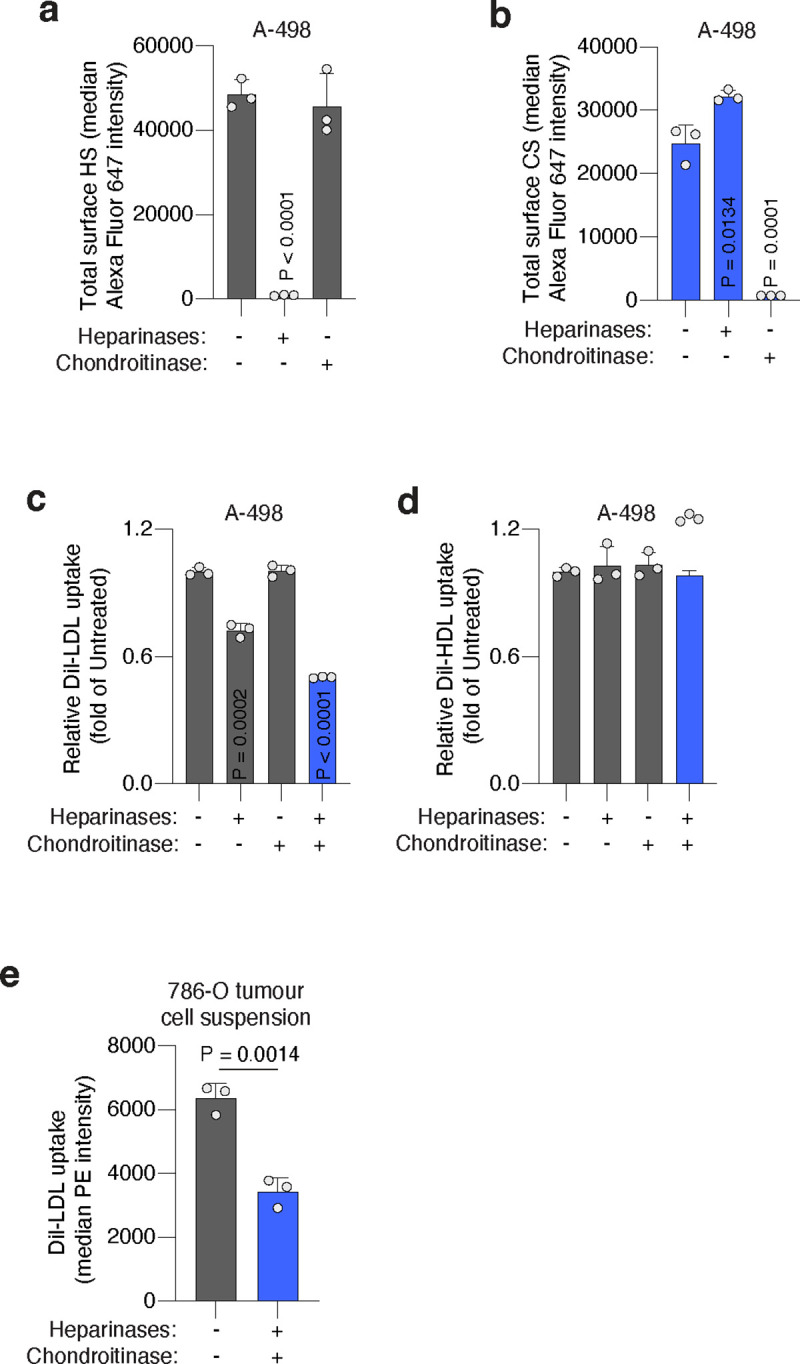
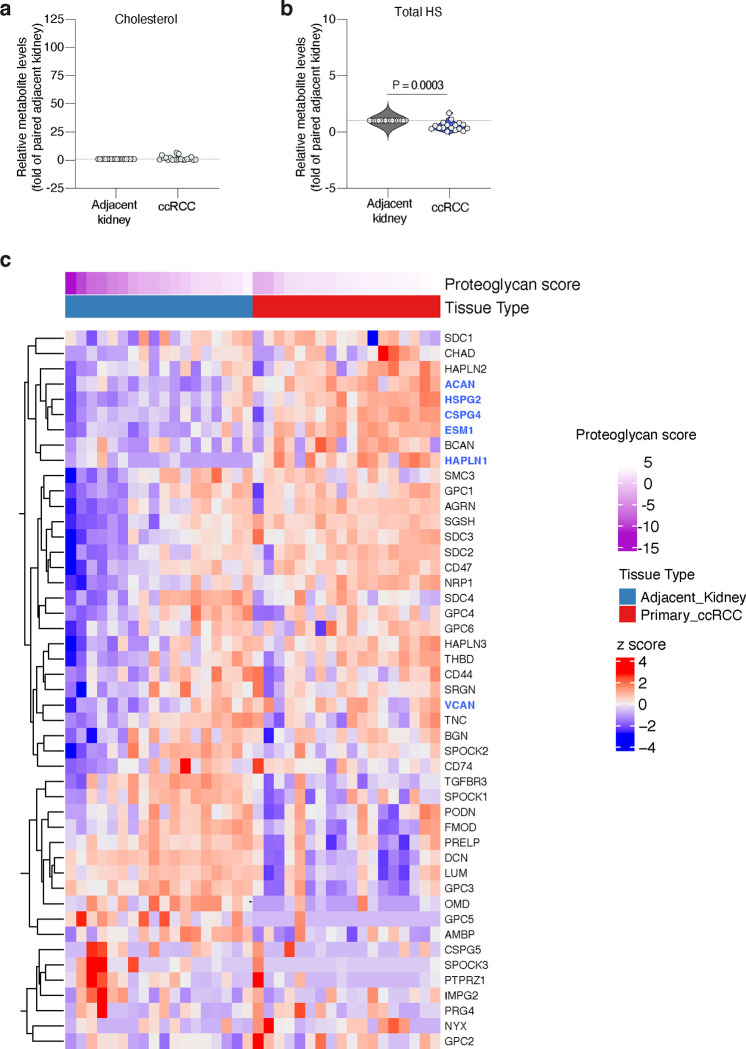
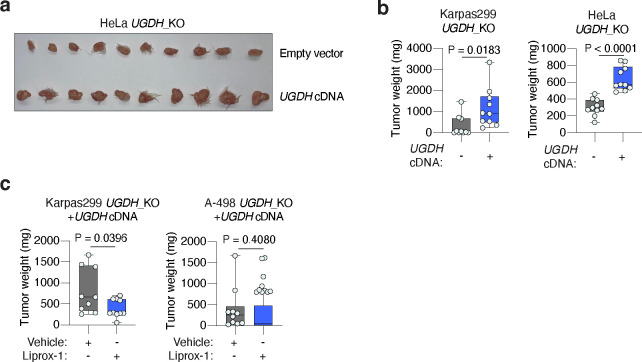
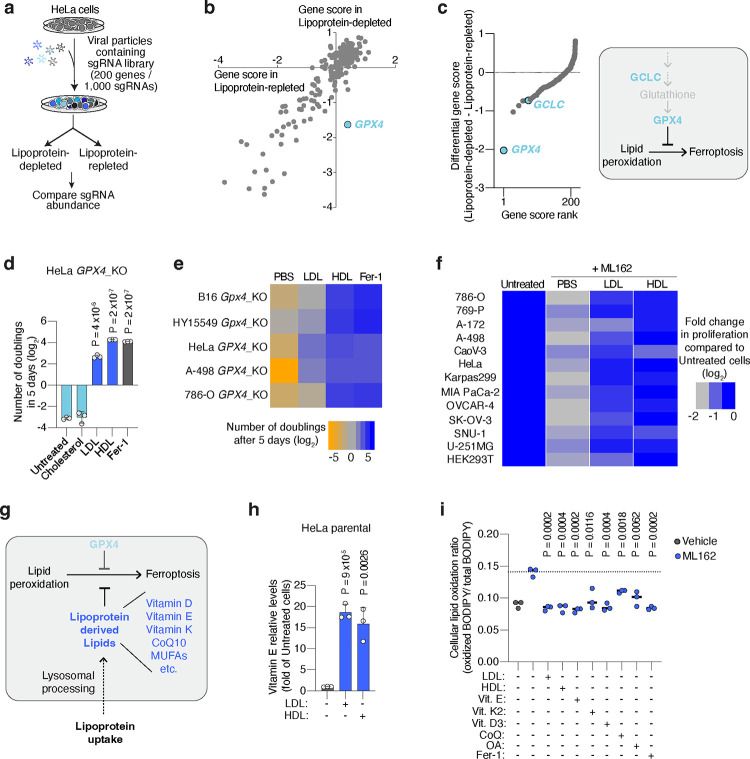
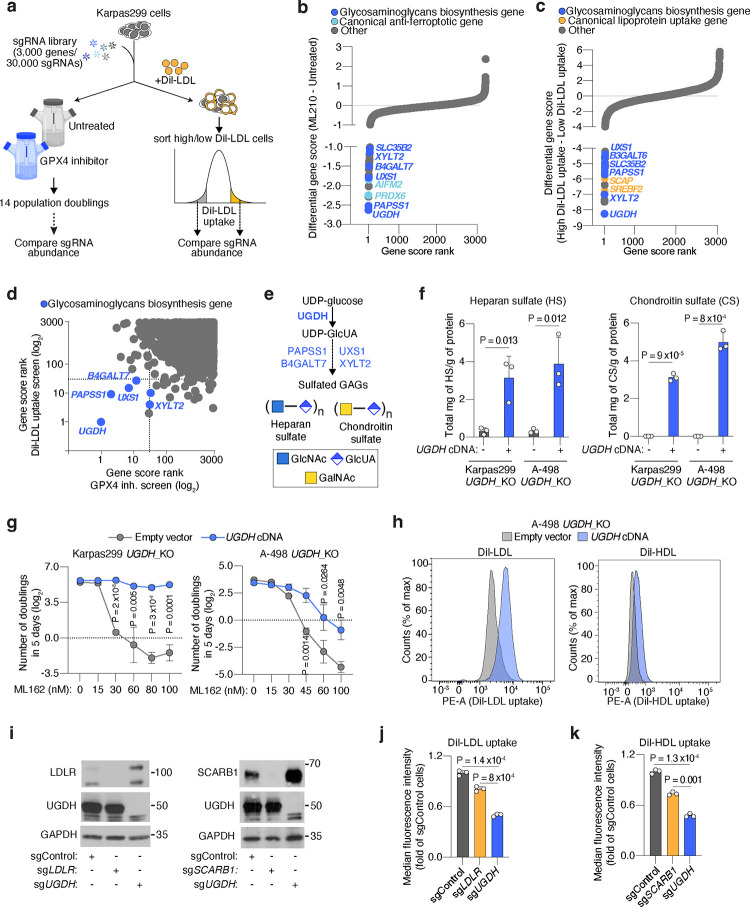
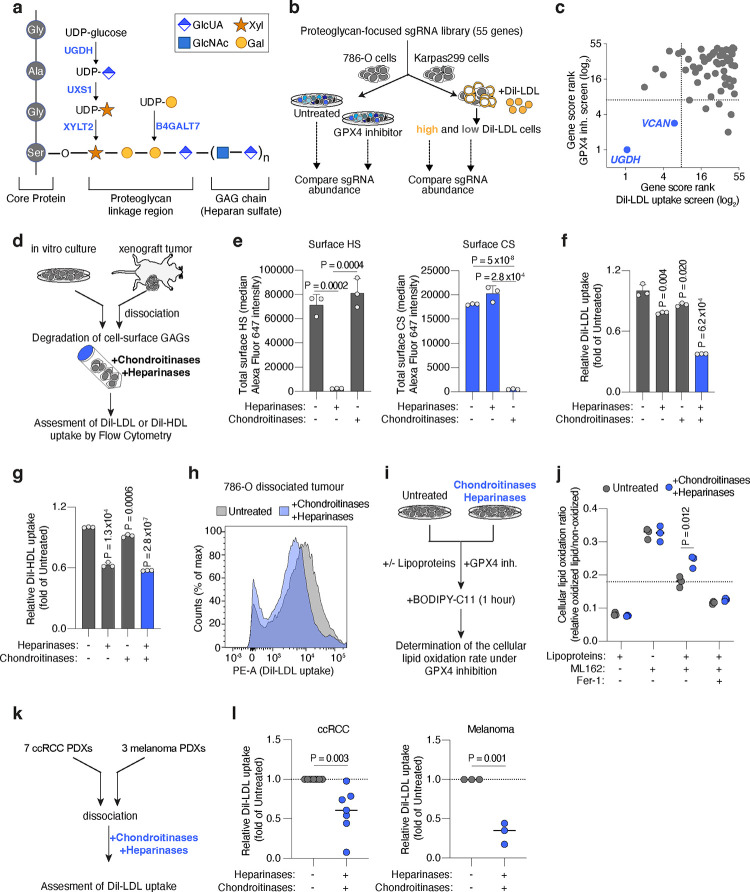
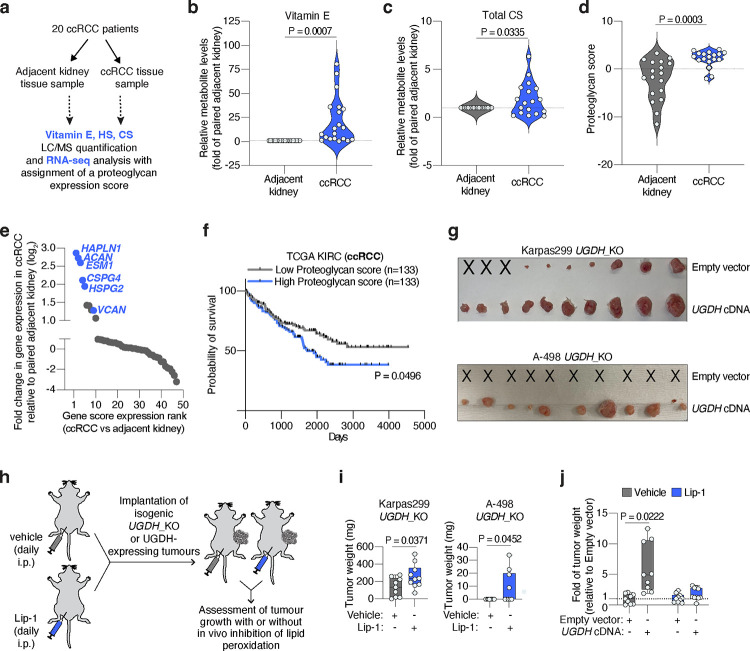
Lipids are essential for tumours because of their structural, energetic, and signaling roles. While many cancer cells upregulate lipid synthesis, growing evidence suggests that tumours simultaneously intensify the uptake of circulating lipids carried by lipoproteins. Which mechanisms promote the uptake of extracellular lipids, and how this pool of lipids contributes to cancer progression, are poorly understood. Here, using functional genetic screens, we find that lipoprotein uptake confers resistance to lipid peroxidation and ferroptotic cell death. Lipoprotein supplementation robustly inhibits ferroptosis across numerous cancer types. Mechanistically, cancer cells take up lipoproteins through a pathway dependent on sulfated glycosaminoglycans (GAGs) linked to cell-surface proteoglycans. Tumour GAGs are a major determinant of the uptake of both low and high density lipoproteins. Impairment of glycosaminoglycan synthesis or acute degradation of surface GAGs decreases the uptake of lipoproteins, sensitizes cells to ferroptosis and reduces tumour growth in mice. We also find that human clear cell renal cell carcinomas, a distinctively lipid-rich tumour type, display elevated levels of lipoprotein-derived antioxidants and the GAG chondroitin sulfate than non-malignant human kidney. Altogether, our work identifies lipoprotein uptake as an essential anti-ferroptotic mechanism for cancer cells to overcome lipid oxidative stress in vivo, and reveals GAG biosynthesis as an unexpected mediator of this process.
Conflict of interest statement
DECLARATION OF INTERESTS K.B. is scientific advisor to Nanocare Pharmaceuticals and Atavistik Bio. R.J.D. is a founder at Atavistik Bio and serves on the Scientific Advisory Boards of Atavistik Bio, Agios Pharmaceuticals, Faeth Therapeutics, General Metabolics and Vida Ventures.
Figures
References
-
- Medes G., Thomas A. & Weinhouse S. Metabolism of neoplastic tissue. IV. A study of lipid synthesis in neoplastic tissue slices in vitro. Cancer Res 13, 27–29 (1953). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources