

This is a preprint.
High dose dietary vitamin D allocates surplus calories to muscle and growth instead of fat via modulation of myostatin and leptin signaling
- PMID: 38766160
- PMCID: PMC11100886
- DOI: 10.21203/rs.3.rs-4202165/v1
High dose dietary vitamin D allocates surplus calories to muscle and growth instead of fat via modulation of myostatin and leptin signaling
Abstract
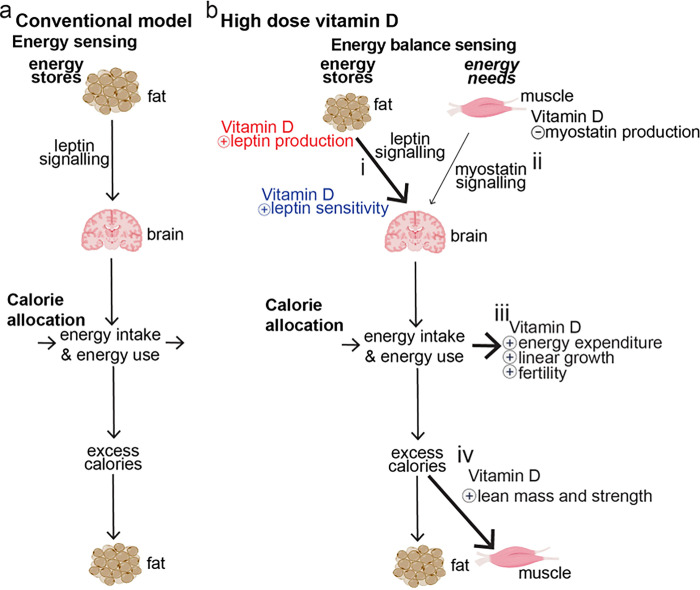
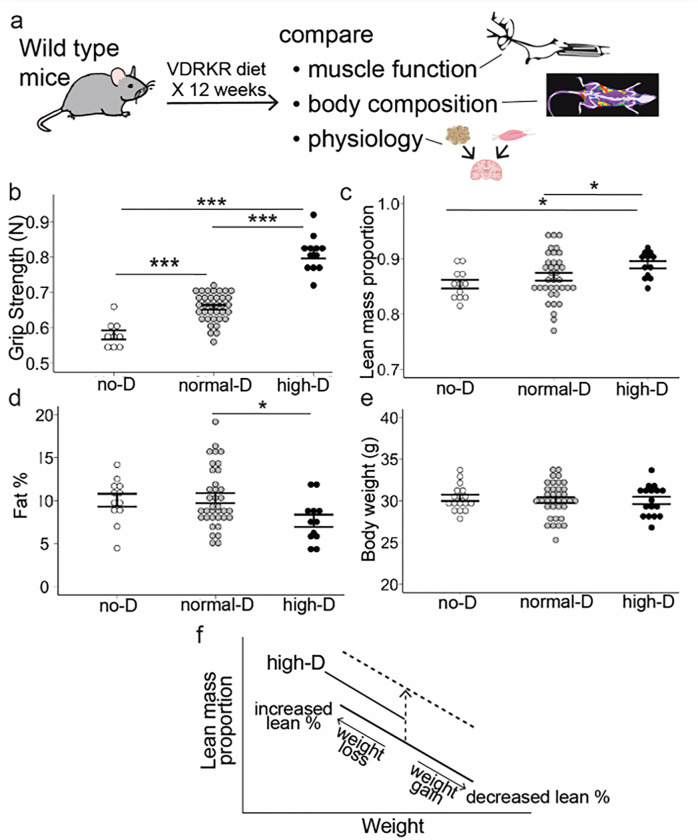
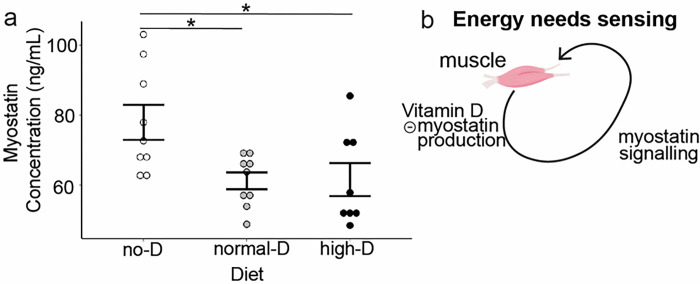
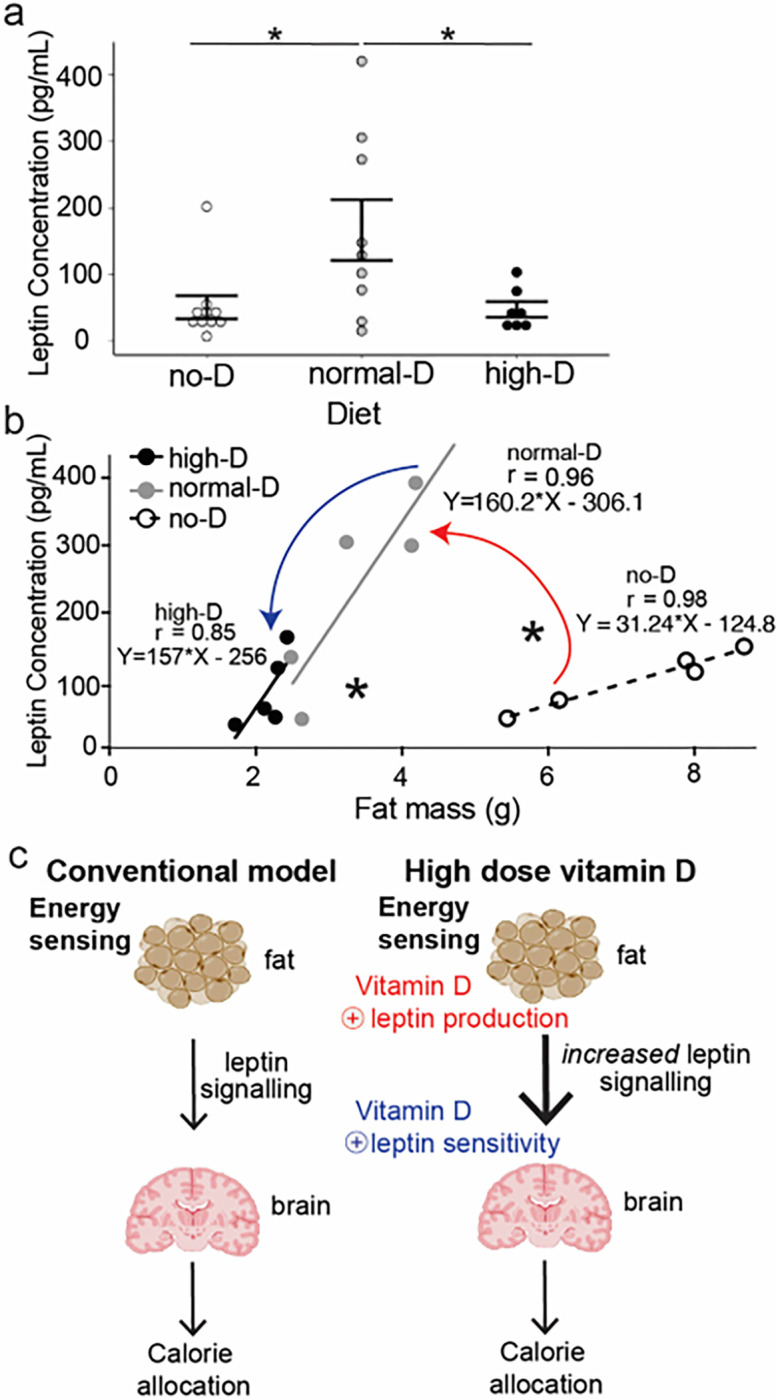
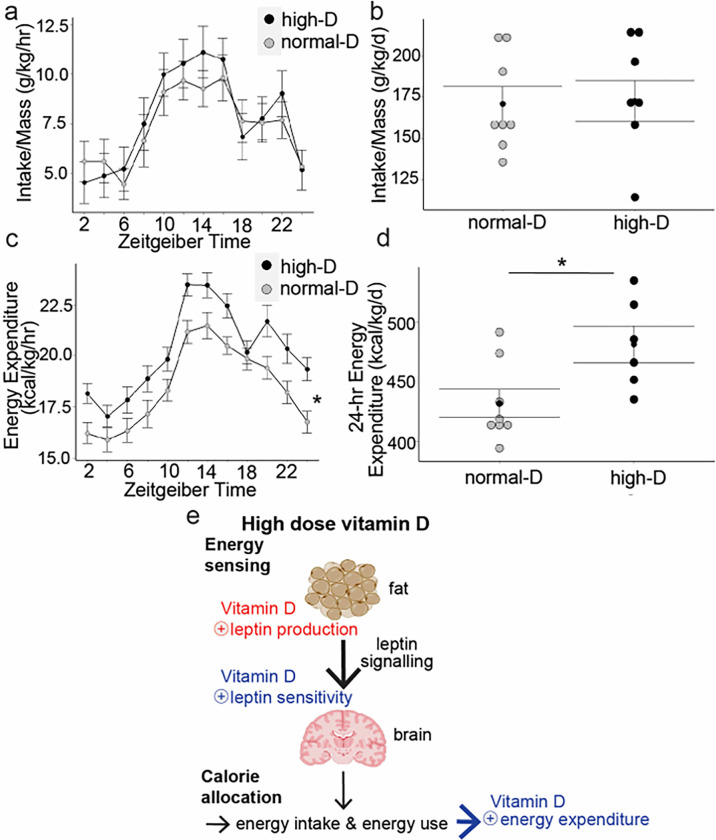
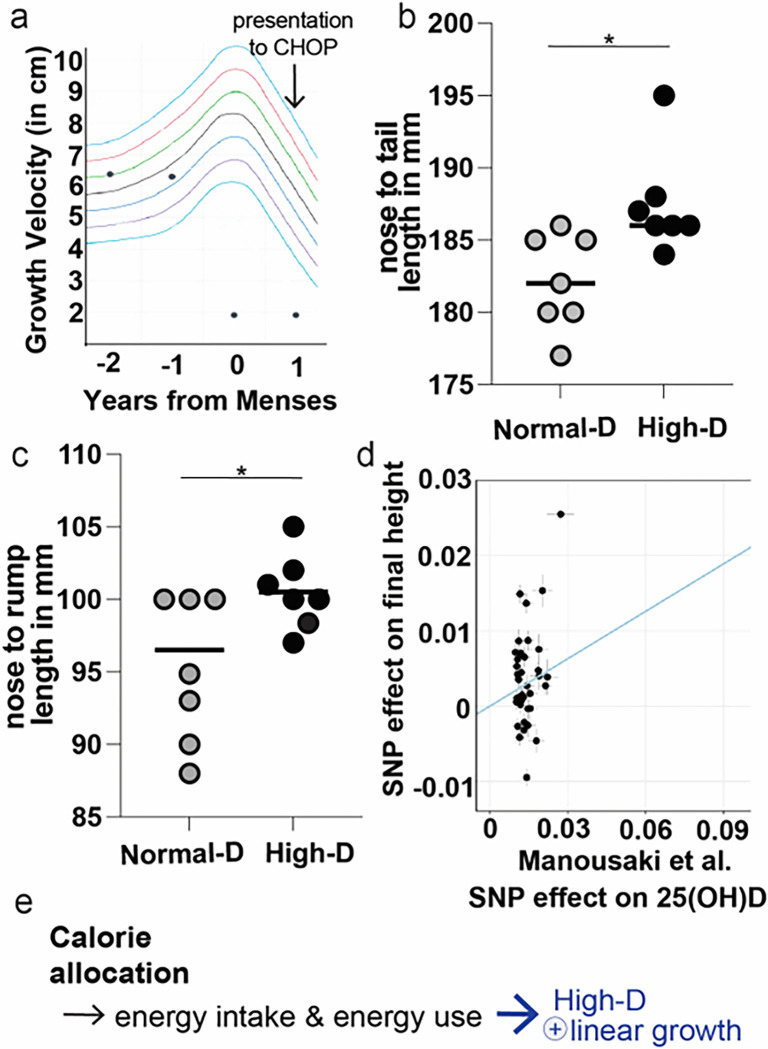
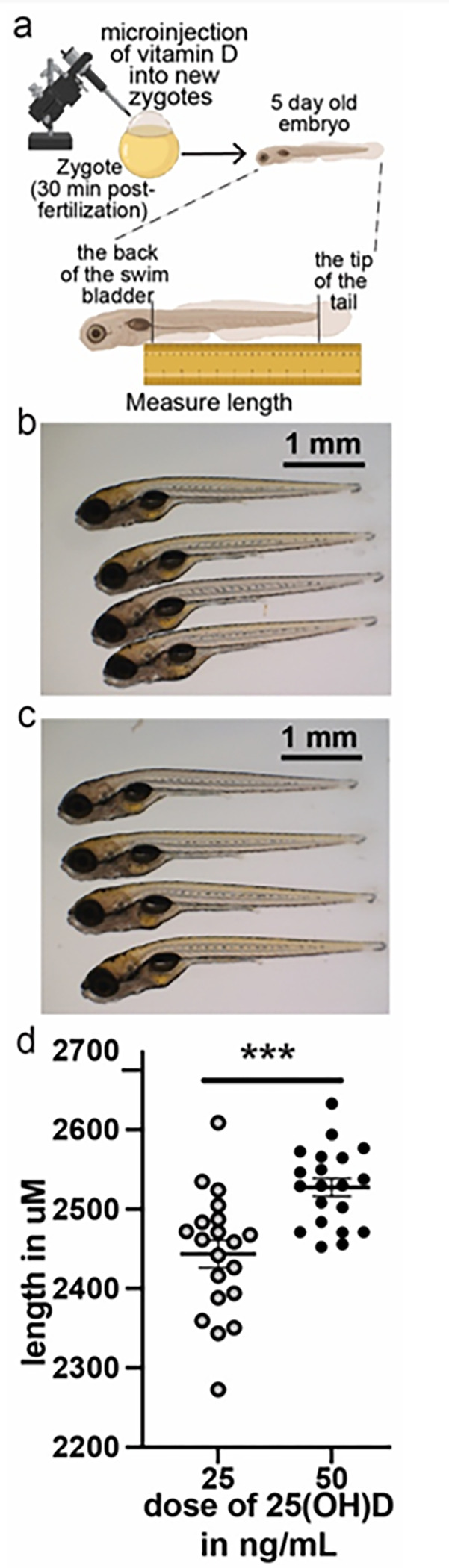
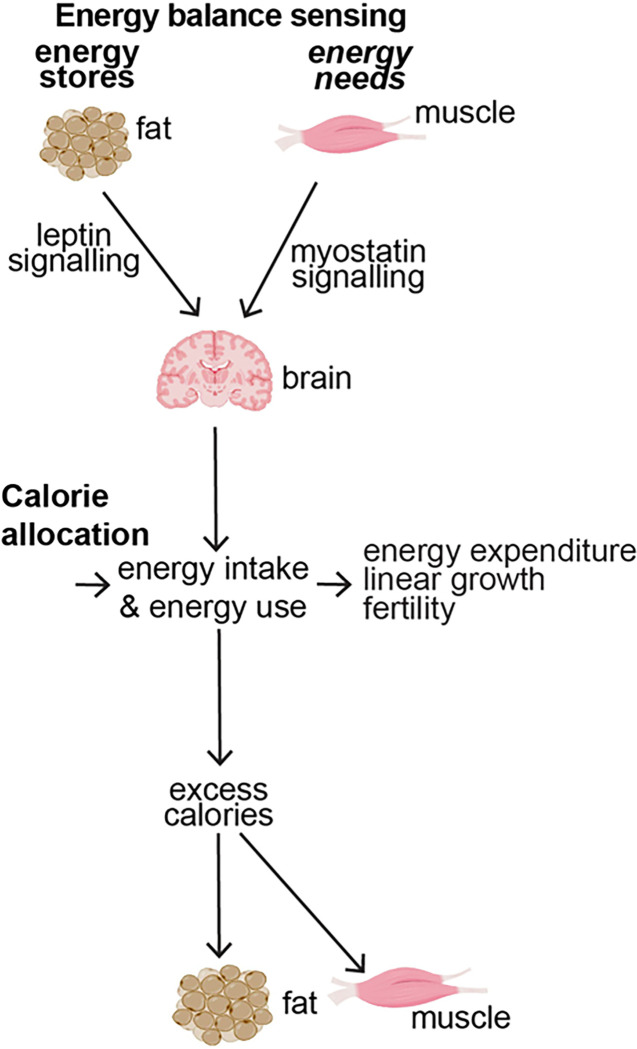
Obesity occurs because the body stores surplus calories as fat rather than as muscle. Fat secretes a hormone, leptin, that modulates energy balance at the brain. Changes in fat mass are mirrored by changes in serum leptin. Elevated leptin prompts the brain to decrease appetite and increase energy expenditure. In obesity, however, impaired leptin sensitivity mutes these leptin-mediated changes. We have limited understanding of what controls leptin production by fat or leptin sensitivity in the brain. Muscle produces a hormone, myostatin, that plays a role in muscle analogous to the one that leptin plays in fat. Absent myostatin leads to increased muscle mass and strength. As with leptin, we also do not know what controls myostatin production or sensitivity. Although fat mass and muscle mass are closely linked, the interplay between leptin and myostatin remains obscure. Here we describe an interplay linked thru vitamin D. Conventionally, it is thought that vitamin D improves strength via trophic effects at the muscle. However, we find here that high dose dietary vitamin D allocates excess calories to muscle and linear growth instead of storage as fat. Vitamin D mediates this allocation by decreasing myostatin production and increasing leptin production and sensitivity. That is, high dose vitamin D improves integration of organismal energy balance. Obesity, aging and other chronic inflammatory diseases are associated with increased fat mass and decreased muscle mass and function (e.g. sarcopenia). Our work provides a physiologic framework for how high-dose vitamin D would increase allocation of calories to muscle instead of fat in these pathologies. Additionally, our work reveals a novel link between the myostatin and leptin signaling whereby myostatin conveys energy needs to modulate leptin effects on calorie allocation. This result provides evidence to update the conventional model of energy stores sensing to a new model of energy balance sensing. In our proposed model, integration of leptin and myostatin signaling allows control of body composition independent of weight. Furthermore, our work reveals how physiologic seasonal variation in vitamin D may be important in controlling season-specific metabolism and calorie allocation to fat in winter and muscle and growth in summer.
Keywords: calorie allocation; energy sensing; growth; leptin; metabolism; myostatin; seasonal; vitamin D.
Conflict of interest statement
Conflict of Interest Statement: The authors have declared that no conflict of interest exists.
Figures
References
-
- Kong J, Chen Y, Zhu G, Zhao Q, Li YC (2013) 1,25-Dihydroxyvitamin D3 upregulates leptin expression in mouse adipose tissue. J Endocrinol 216:265–271 - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
