

This is a preprint.
Pharmacological inhibition of tyrosine protein-kinase 2 reduces islet inflammation and delays type 1 diabetes onset in mice
- PMID: 38766166
- PMCID: PMC11100605
- DOI: 10.1101/2024.03.20.585925
Pharmacological inhibition of tyrosine protein-kinase 2 reduces islet inflammation and delays type 1 diabetes onset in mice
Update in
-
Pharmacological inhibition of tyrosine protein-kinase 2 reduces islet inflammation and delays type 1 diabetes onset in mice.EBioMedicine. 2025 Jul;117:105734. doi: 10.1016/j.ebiom.2025.105734. Epub 2025 May 6. EBioMedicine. 2025. PMID: 40335415 Free PMC article.
Abstract
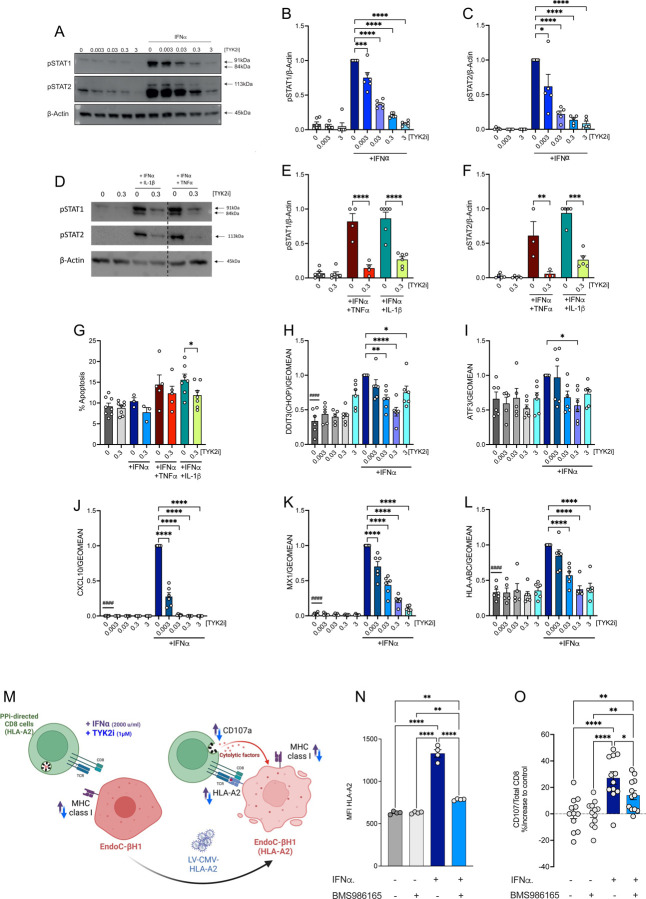
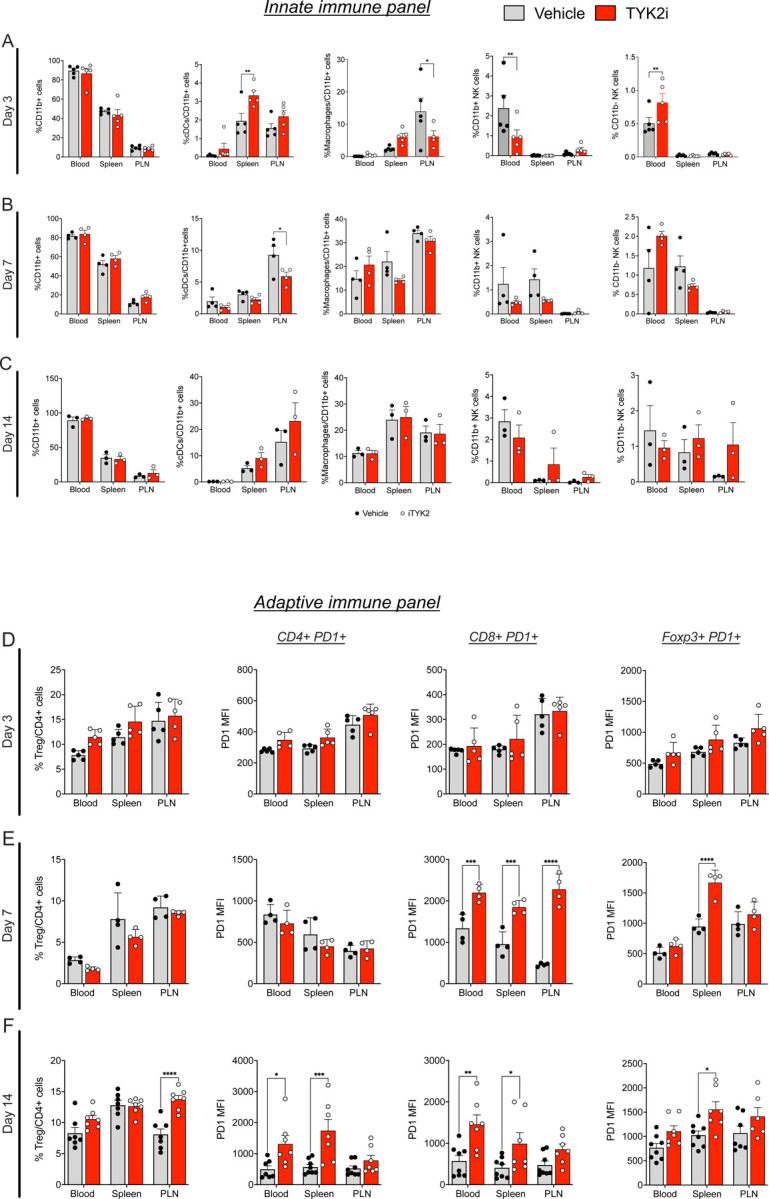
Tyrosine protein-kinase 2 (TYK2), a member of the Janus kinase family, mediates inflammatory signaling through multiple cytokines, including interferon-α (IFNα), interleukin (IL)-12, and IL-23. Missense mutations in TYK2 are associated with protection against type 1 diabetes (T1D), and inhibition of TYK2 shows promise in the management of other autoimmune conditions. Here, we evaluated the effects of specific TYK2 inhibitors (TYK2is) in pre-clinical models of T1D. First, human β cells, cadaveric donor islets, and iPSC-derived islets were treated in vitro with IFNα in combination with a small molecule TYK2i (BMS-986165 or a related molecule BMS-986202). TYK2 inhibition prevented IFNα-induced β cell HLA class I up-regulation, endoplasmic reticulum stress, and chemokine production. In co-culture studies, pre-treatment of β cells with a TYK2i prevented IFNα-induced activation of T cells targeting an epitope of insulin. In vivo administration of BMS-986202 in two mouse models of T1D (RIP-LCMV-GP mice and NOD mice) reduced systemic and tissue-localized inflammation, prevented β cell death, and delayed T1D onset. Transcriptional phenotyping of pancreatic islets, pancreatic lymph nodes (PLN), and spleen during early disease pathogenesis highlighted a role for TYK2 inhibition in modulating signaling pathways associated with inflammation, translational control, stress signaling, secretory function, immunity, and diabetes. Additionally, TYK2i treatment changed the composition of innate and adaptive immune cell populations in the blood and disease target tissues, resulting in an immune phenotype with a diminished capacity for β cell destruction. Overall, these findings indicate that TYK2i has beneficial effects in both the immune and endocrine compartments in models of T1D, thus supporting a path forward for testing TYK2 inhibitors in human T1D.
Keywords: T cell; Type 1 diabetes; interferon-α; islets of Langerhans; tyrosine protein-kinase 2 (TYK2); β cell.
Conflict of interest statement
Declaration of Competing Interest: CEM has served on advisory boards related to T1D research clinical trial initiatives: Provention Bio, Dompe Pharmaceuticals, Isla Technologies, MaiCell Technologies, Avotres, and DiogenX. CEM has patent (16/291,668) Extracellular Vesicle Ribonucleic Acid (RNA) Cargo as a Biomarker of Hyperglycaemia and Type 1 Diabetes and CEM and FS have a provisional patent (63/285,765) Biomarker for Type 1 Diabetes (PDIA1 as a biomarker of β cell stress). These activities have not dealt directly with topics covered in this manuscript.
Figures






References
-
- Platanias LC. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat Rev Immunol. 2005;5(5):375–86. - PubMed
-
- Tao JH, Zou YF, Feng XL, Li J, Wang F, Pan FM, et al. Meta-analysis of TYK2 gene polymorphisms association with susceptibility to autoimmune and inflammatory diseases. Mol Biol Rep. 2011;38(7):4663–72. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous