

Targeting chondroitin sulfate suppresses macropinocytosis of breast cancer cells by modulating syndecan-1 expression
- PMID: 38770553
- PMCID: PMC11459036
- DOI: 10.1002/1878-0261.13667
Targeting chondroitin sulfate suppresses macropinocytosis of breast cancer cells by modulating syndecan-1 expression
Abstract
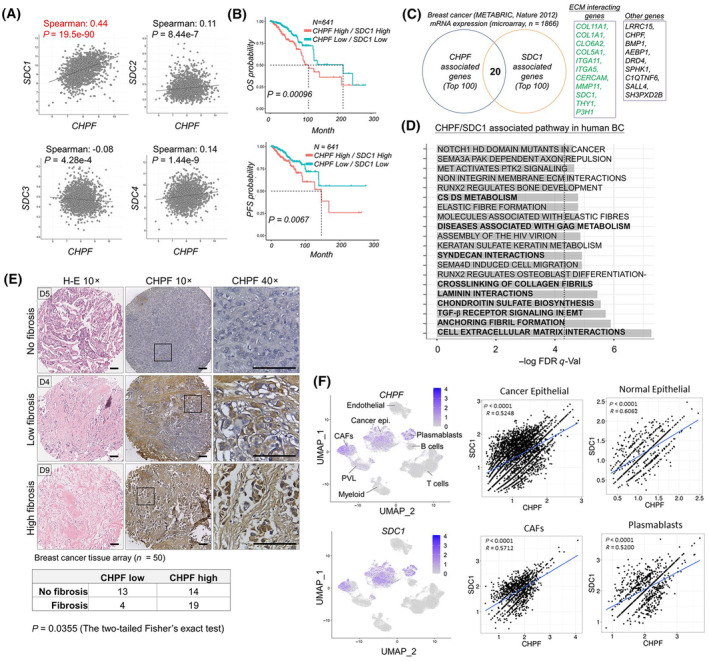
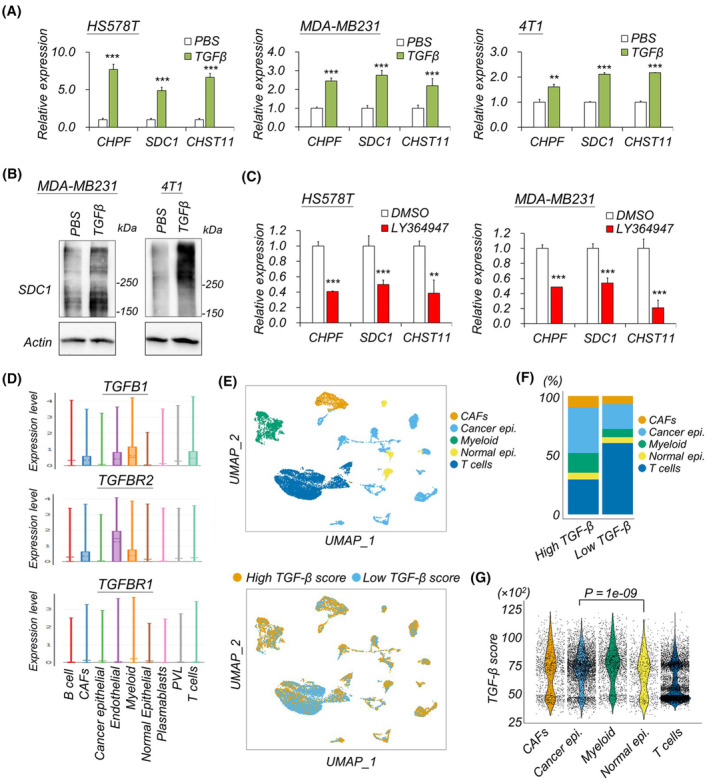
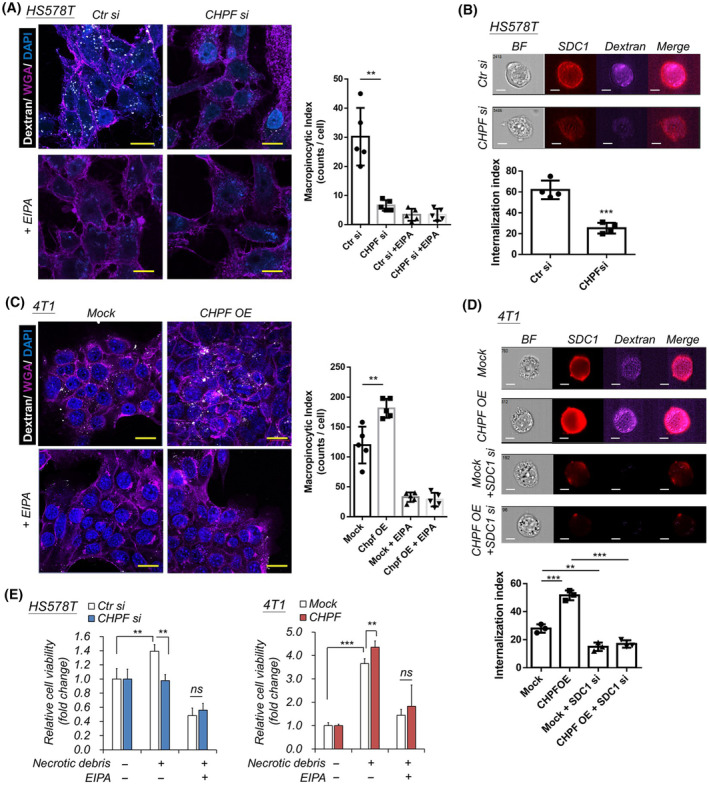
Accumulation of abnormal chondroitin sulfate (CS) chains in breast cancer tissue is correlated with poor prognosis. However, the biological functions of these CS chains in cancer progression remain largely unknown, impeding the development of targeted treatment focused on CS. Previous studies identified chondroitin polymerizing factor (CHPF; also known as chondroitin sulfate synthase 2) is the critical enzyme regulating CS accumulation in breast cancer tissue. We then assessed the association between CHPF-associated proteoglycans (PGs) and signaling pathways in breast cancer datasets. The regulation between CHPF and syndecan 1 (SDC1) was examined at both the protein and RNA levels. Confocal microscopy and image flow cytometry were employed to quantify macropinocytosis. The effects of the 6-O-sulfated CS-binding peptide (C6S-p) on blocking CS functions were tested in vitro and in vivo. Results indicated that the expression of CHPF and SDC1 was tightly associated within primary breast cancer tissue, and high expression of both genes exacerbated patient prognosis. Transforming growth factor beta (TGF-β) signaling was implicated in the regulation of CHPF and SDC1 in breast cancer cells. CHPF supported CS-SDC1 stabilization on the cell surface, modulating macropinocytotic activity in breast cancer cells under nutrient-deprived conditions. Furthermore, C6S-p demonstrated the ability to bind CS-SDC1, increase SDC1 degradation, suppress macropinocytosis of breast cancer cells, and inhibit tumor growth in vivo. Although other PGs may also be involved in CHPF-regulated breast cancer malignancy, this study provides the first evidence that a CS synthase participates in the regulation of macropinocytosis in cancer cells by supporting SDC1 expression on cancer cells.
Keywords: CHPF; breast cancer; chondroitin sulfate; macropinocytosis; syndecan 1.
© 2024 The Authors. Molecular Oncology published by John Wiley & Sons Ltd on behalf of Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Neill T, Painter H, Buraschi S, Owens RT, Lisanti MP, Schaefer L, et al. Decorin antagonizes the angiogenic network: concurrent inhibition of Met, hypoxia inducible factor 1alpha, vascular endothelial growth factor A, and induction of thrombospondin‐1 and TIMP3. J Biol Chem. 2012;287:5492–5506. 10.1074/jbc.M111.283499 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
