

Ciliary tip actin dynamics regulate photoreceptor outer segment integrity
- PMID: 38773095
- PMCID: PMC11109262
- DOI: 10.1038/s41467-024-48639-w
Ciliary tip actin dynamics regulate photoreceptor outer segment integrity
Abstract
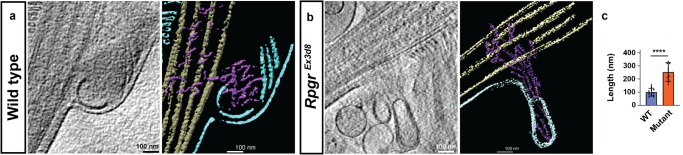
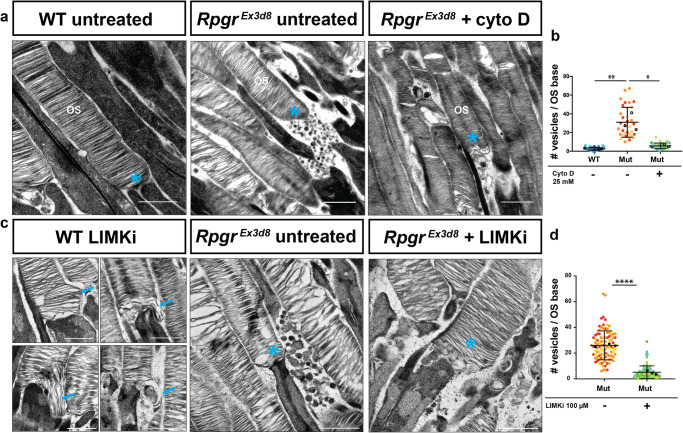
As signalling organelles, cilia regulate their G protein-coupled receptor content by ectocytosis, a process requiring localised actin dynamics to alter membrane shape. Photoreceptor outer segments comprise an expanse of folded membranes (discs) at the tip of highly-specialised connecting cilia, into which photosensitive GPCRs are concentrated. Discs are shed and remade daily. Defects in this process, due to mutations, cause retinitis pigmentosa (RP). Whilst fundamental for vision, the mechanism of photoreceptor disc generation is poorly understood. Here, we show membrane deformation required for disc genesis is driven by dynamic actin changes in a process akin to ectocytosis. We show RPGR, a leading RP gene, regulates actin-binding protein activity central to this process. Actin dynamics, required for disc formation, are perturbed in Rpgr mouse models, leading to aborted membrane shedding as ectosome-like vesicles, photoreceptor death and visual loss. Actin manipulation partially rescues this, suggesting the pathway could be targeted therapeutically. These findings help define how actin-mediated dynamics control outer segment turnover.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
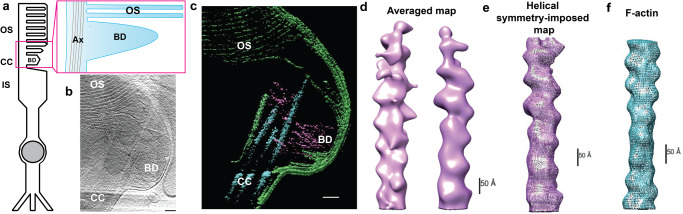
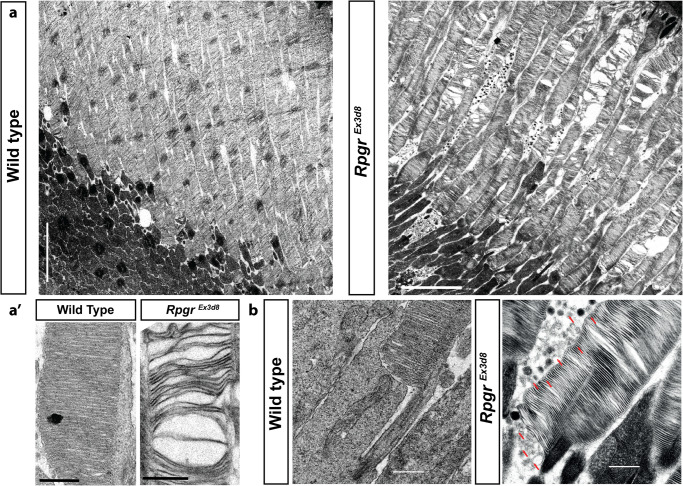
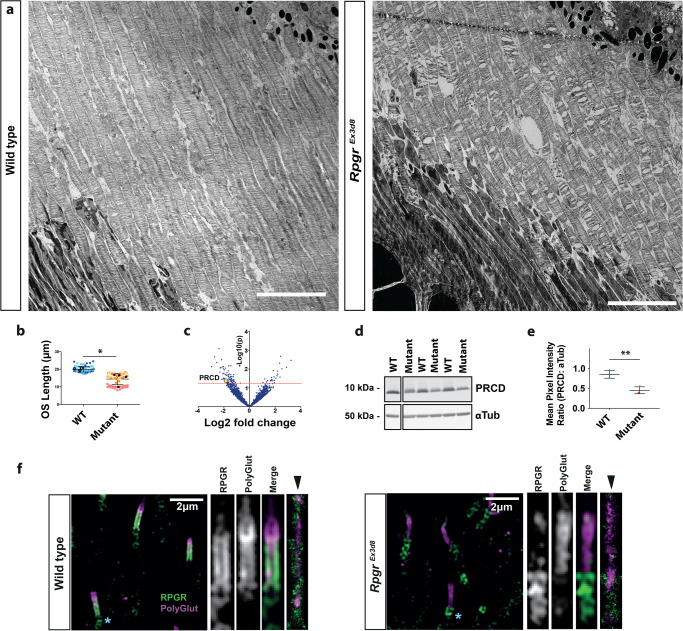
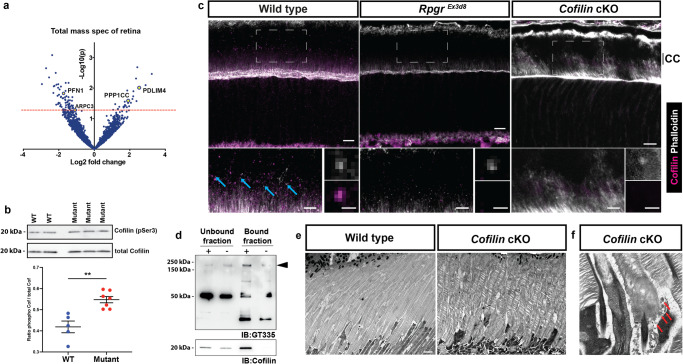
References
-
- Papermaster DS, Schneider BG, Besharse JC. Vesicular transport of newly synthesized opsin from the Golgi apparatus toward the rod outer segment. Ultrastructural immunocytochemical and autoradiographic evidence in Xenopus retinas. Invest. Ophthalmol. Vis. Sci. 1985;26:1386–1404. - PubMed
MeSH terms
Grants and funding
- MC_UU_00007_14/RCUK | Medical Research Council (MRC)
- Q-0035/Welch Foundation
- 5179 / 5180/Fight for Sight UK
- R01 EY030096/EY/NEI NIH HHS/United States
- MR_Y015002_1/RCUK | Medical Research Council (MRC)
- R01 EY026545/EY/NEI NIH HHS/United States
- MC_UU_12018/26/RCUK | Medical Research Council (MRC)
- 208402/Z/17/Z/Wellcome Trust (Wellcome)
- WT_/Wellcome Trust/United Kingdom
- MC_UU_00007/14/MRC_/Medical Research Council/United Kingdom
- MC_PC_21044/RCUK | Medical Research Council (MRC)
- A24452/Cancer Research UK (CRUK)
- R01 EY031949/EY/NEI NIH HHS/United States
- R01-EY026545/U.S. Department of Health & Human Services | NIH | National Eye Institute (NEI)
- F32 EY031574/EY/NEI NIH HHS/United States
- R01-EY026545 and R01-EY031949/U.S. Department of Health & Human Services | NIH | National Eye Institute (NEI)
- R01-EY031949/U.S. Department of Health & Human Services | NIH | National Eye Institute (NEI)
- 215343/Z/19/Z/Wellcome Trust (Wellcome)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
