

Diurnal control of iron responsive element containing mRNAs through iron regulatory proteins IRP1 and IRP2 is mediated by feeding rhythms
- PMID: 38773499
- PMCID: PMC11106963
- DOI: 10.1186/s13059-024-03270-2
Diurnal control of iron responsive element containing mRNAs through iron regulatory proteins IRP1 and IRP2 is mediated by feeding rhythms
Abstract
Background: Cellular iron homeostasis is regulated by iron regulatory proteins (IRP1 and IRP2) that sense iron levels (and other metabolic cues) and modulate mRNA translation or stability via interaction with iron regulatory elements (IREs). IRP2 is viewed as the primary regulator in the liver, yet our previous datasets showing diurnal rhythms for certain IRE-containing mRNAs suggest a nuanced temporal control mechanism. The purpose of this study is to gain insights into the daily regulatory dynamics across IRE-bearing mRNAs, specific IRP involvement, and underlying systemic and cellular rhythmicity cues in mouse liver.
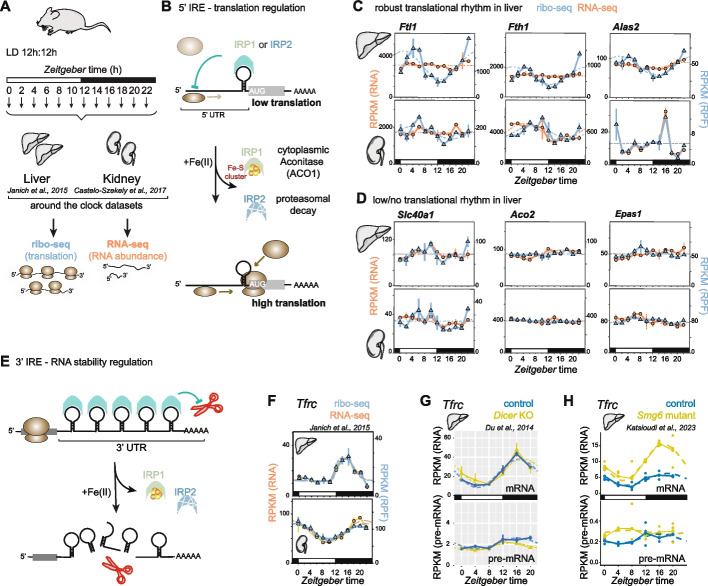
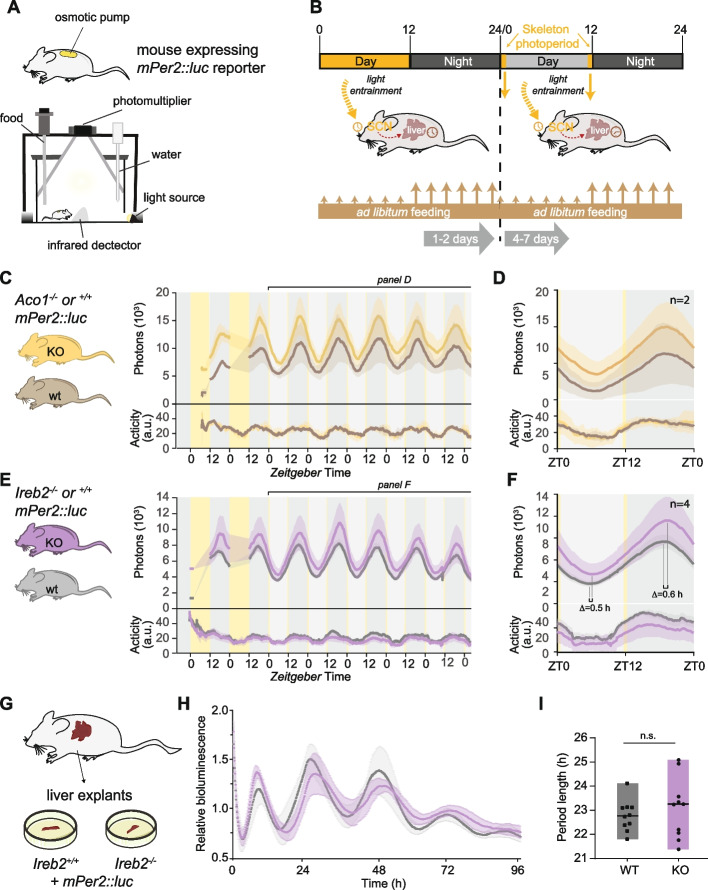
Results: We uncover high-amplitude diurnal oscillations in the regulation of key IRE-containing transcripts in the liver, compatible with maximal IRP activity at the onset of the dark phase. Although IRP2 protein levels also exhibit some diurnal variations and peak at the light-dark transition, ribosome profiling in IRP2-deficient mice reveals that maximal repression of target mRNAs at this timepoint still occurs. We further find that diurnal regulation of IRE-containing mRNAs can continue in the absence of a functional circadian clock as long as feeding is rhythmic.
Conclusions: Our findings suggest temporally controlled redundancy in IRP activities, with IRP2 mediating regulation of IRE-containing transcripts in the light phase and redundancy, conceivably with IRP1, at dark onset. Moreover, we highlight the significance of feeding-associated signals in driving rhythmicity. Our work highlights the dynamic nature and regulatory complexity in a metabolic pathway that had previously been considered well-understood.
Keywords: Circadian clocks; IRE; IRP; Iron metabolism; Liver; Ribosome profiling; Translation.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
-
- Bolshette N, Ibrahim H, Reinke H, Asher G. Circadian regulation of liver function: from molecular mechanisms to disease pathophysiology. Nat Rev Gastroenterol Hepatol. 2023;20:695–707. - PubMed
-
- Castelo-Szekely V, Gatfield D. Emerging Roles of Translational Control in Circadian Timekeeping. J Mol Biol. 2020;432:3483–3497. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
