

Structure and mechanism of the human CTDNEP1-NEP1R1 membrane protein phosphatase complex necessary to maintain ER membrane morphology
- PMID: 38776370
- PMCID: PMC11145253
- DOI: 10.1073/pnas.2321167121
Structure and mechanism of the human CTDNEP1-NEP1R1 membrane protein phosphatase complex necessary to maintain ER membrane morphology
Abstract
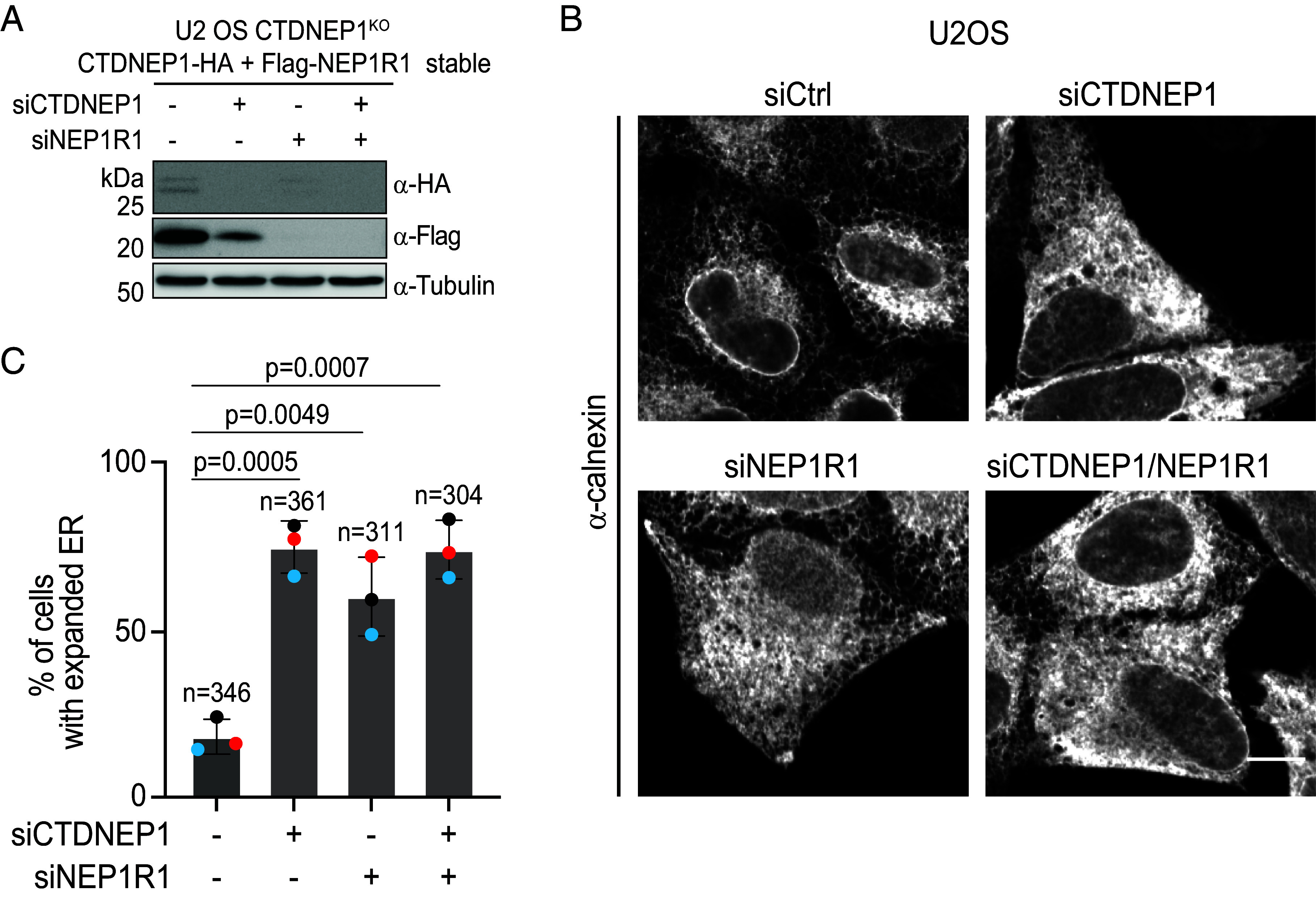
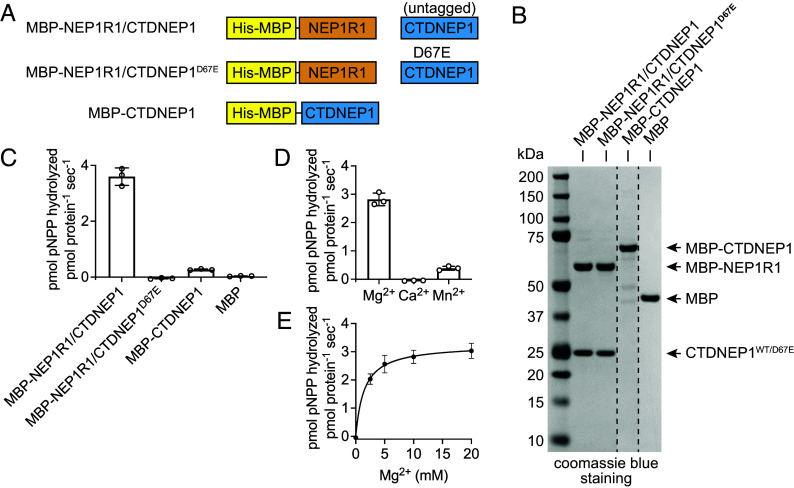
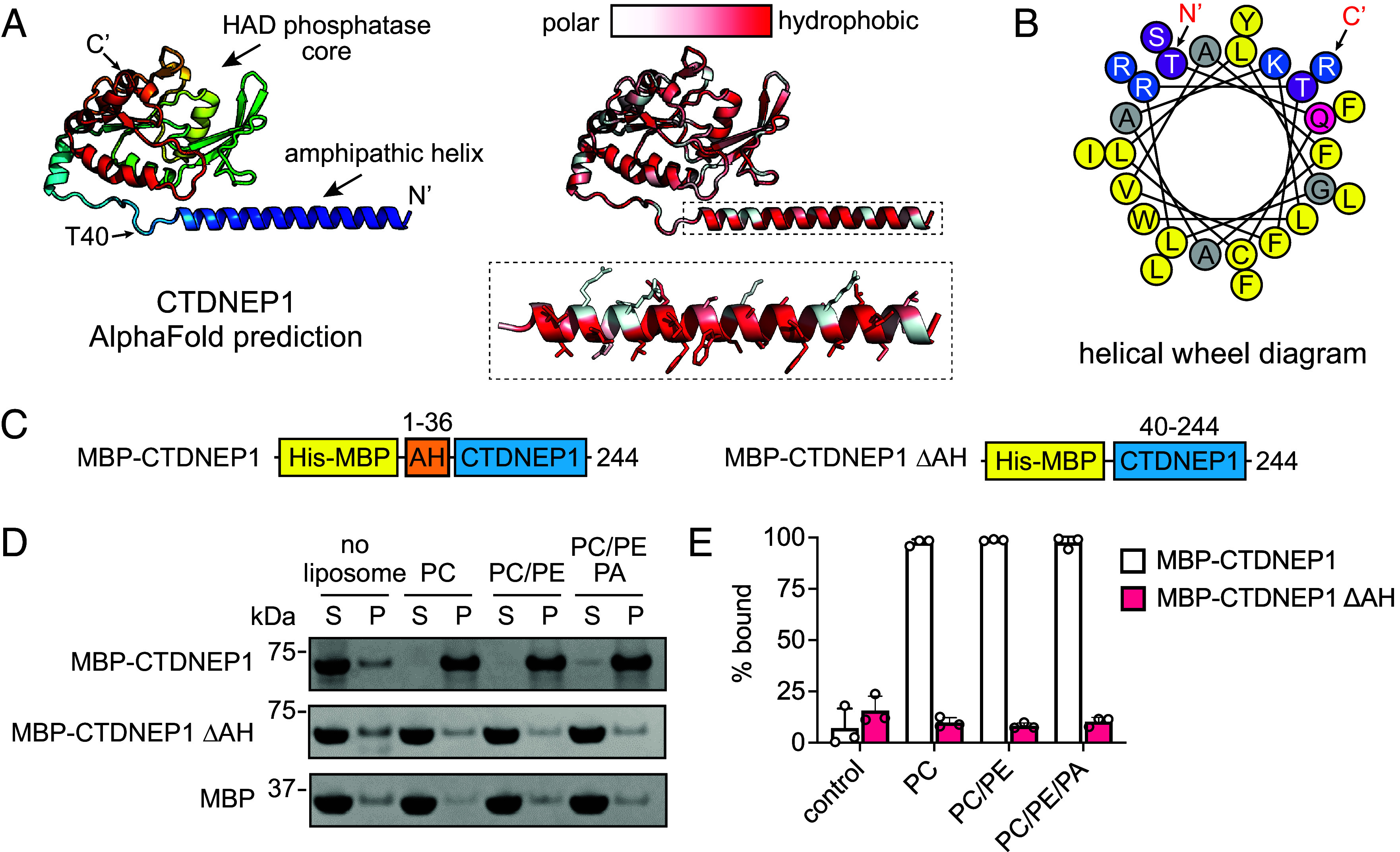
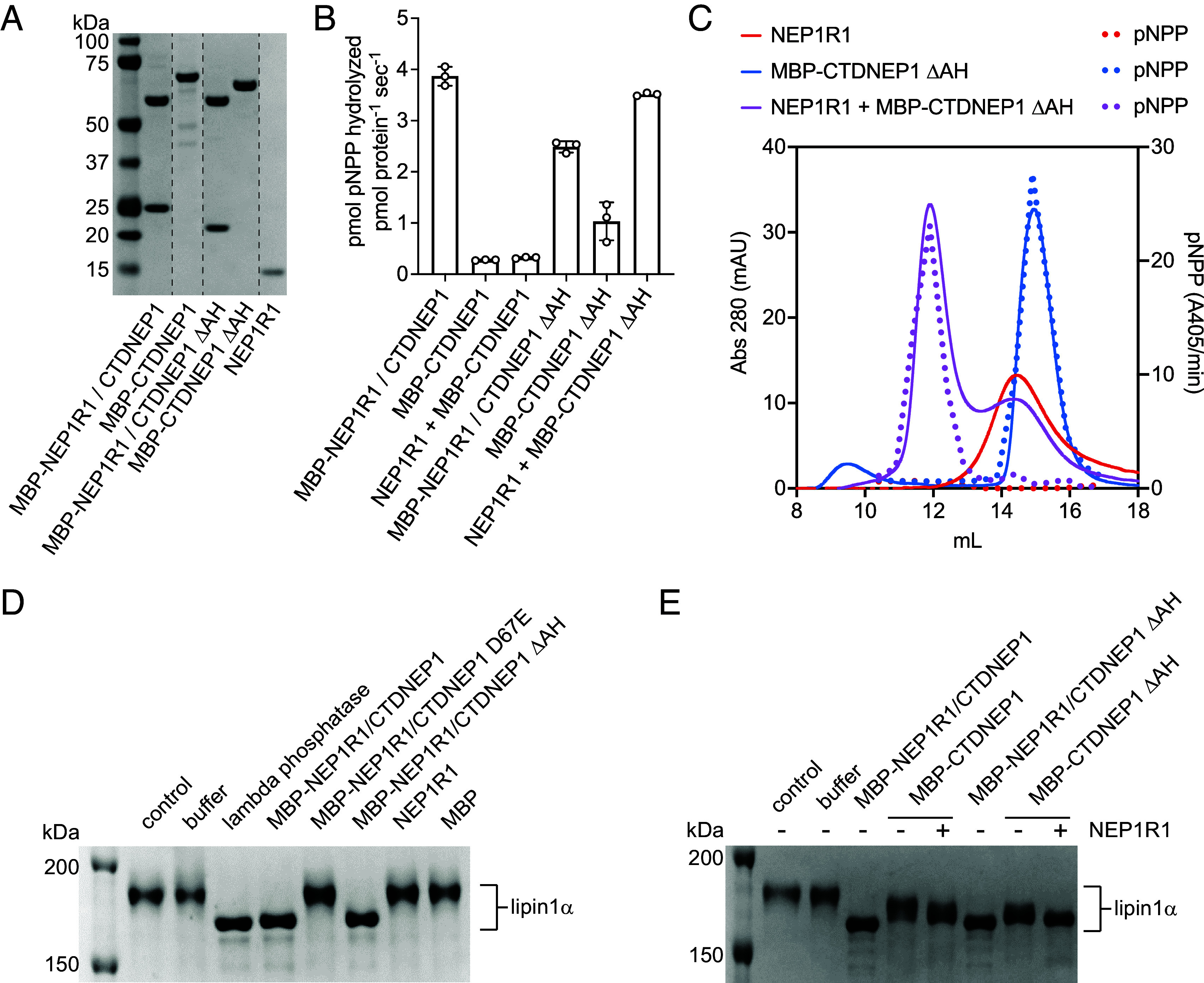
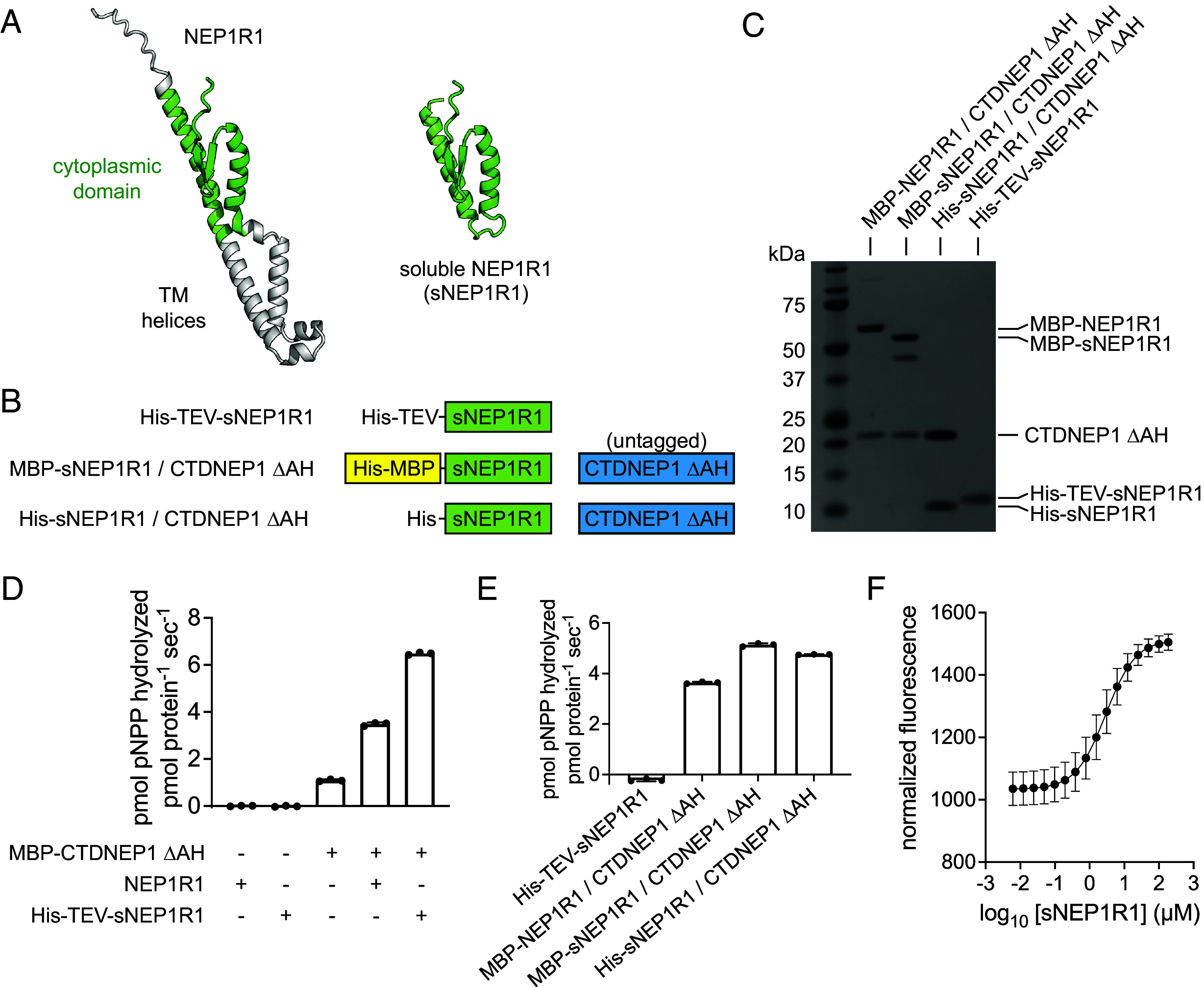
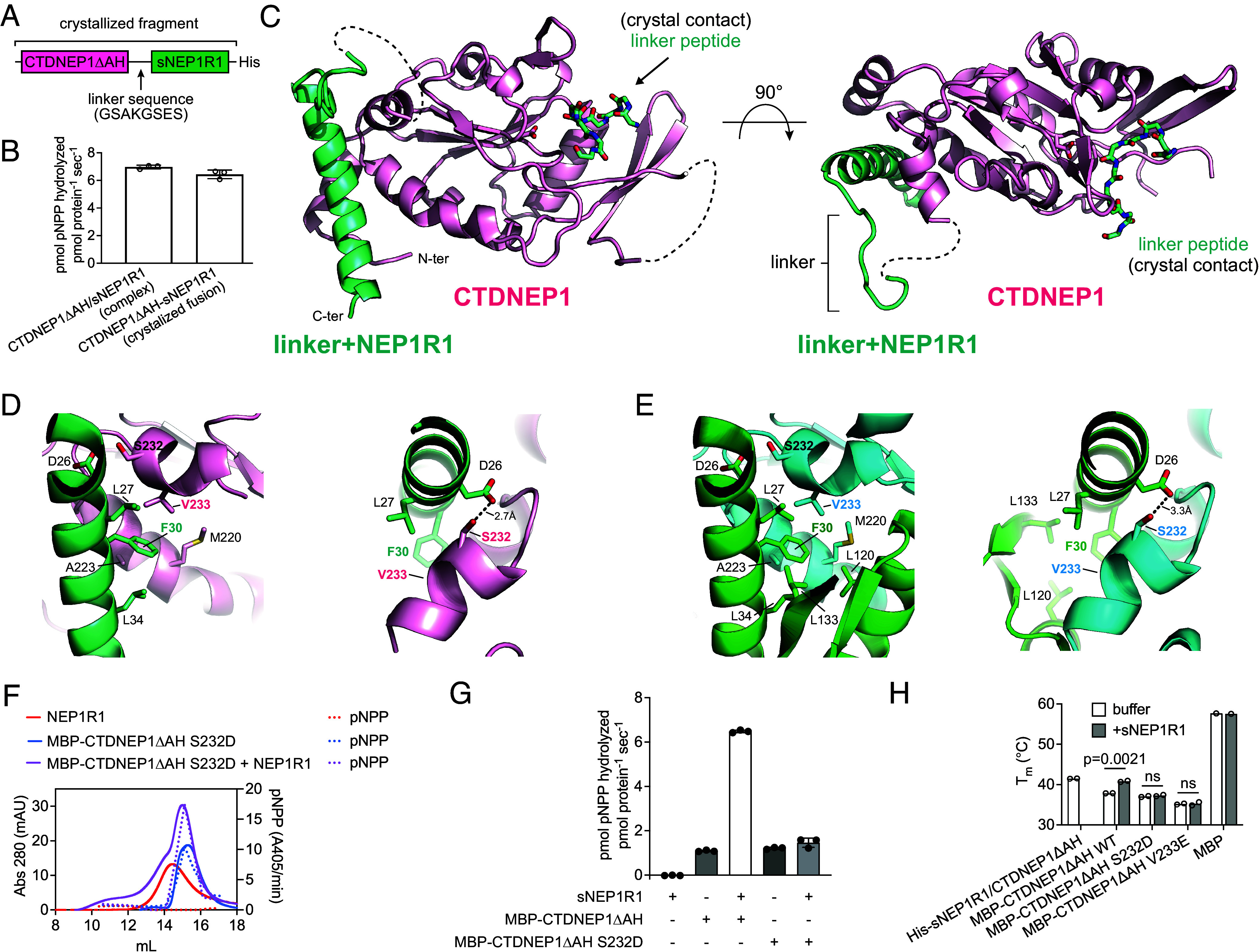
C-terminal Domain Nuclear Envelope Phosphatase 1 (CTDNEP1) is a noncanonical protein serine/threonine phosphatase that has a conserved role in regulating ER membrane biogenesis. Inactivating mutations in CTDNEP1 correlate with the development of medulloblastoma, an aggressive childhood cancer. The transmembrane protein Nuclear Envelope Phosphatase 1 Regulatory Subunit 1 (NEP1R1) binds CTDNEP1, but the molecular details by which NEP1R1 regulates CTDNEP1 function are unclear. Here, we find that knockdown of NEP1R1 generates identical phenotypes to reported loss of CTDNEP1 in mammalian cells, establishing CTDNEP1-NEP1R1 as an evolutionarily conserved membrane protein phosphatase complex that restricts ER expansion. Mechanistically, NEP1R1 acts as an activating regulatory subunit that directly binds and increases the phosphatase activity of CTDNEP1. By defining a minimal NEP1R1 domain sufficient to activate CTDNEP1, we determine high-resolution crystal structures of the CTDNEP1-NEP1R1 complex bound to a peptide sequence acting as a pseudosubstrate. Structurally, NEP1R1 engages CTDNEP1 at a site distant from the active site to stabilize and allosterically activate CTDNEP1. Substrate recognition is facilitated by a conserved Arg residue in CTDNEP1 that binds and orients the substrate peptide in the active site. Together, this reveals mechanisms for how NEP1R1 regulates CTDNEP1 and explains how cancer-associated mutations inactivate CTDNEP1.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures

Update of
-
Structure and mechanism of the human CTDNEP1-NEP1R1 membrane protein phosphatase complex necessary to maintain ER membrane morphology.bioRxiv [Preprint]. 2023 Nov 20:2023.11.20.567952. doi: 10.1101/2023.11.20.567952. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2024 May 28;121(22):e2321167121. doi: 10.1073/pnas.2321167121. PMID: 38045299 Free PMC article. Updated. Preprint.
Similar articles
-
Differential reliance of CTD-nuclear envelope phosphatase 1 on its regulatory subunit in ER lipid synthesis and storage.Mol Biol Cell. 2024 Jul 1;35(7):ar101. doi: 10.1091/mbc.E23-09-0382. Epub 2024 May 22. Mol Biol Cell. 2024. PMID: 38776127 Free PMC article.
-
Structure and mechanism of the human CTDNEP1-NEP1R1 membrane protein phosphatase complex necessary to maintain ER membrane morphology.bioRxiv [Preprint]. 2023 Nov 20:2023.11.20.567952. doi: 10.1101/2023.11.20.567952. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2024 May 28;121(22):e2321167121. doi: 10.1073/pnas.2321167121. PMID: 38045299 Free PMC article. Updated. Preprint.
-
Torsin and NEP1R1-CTDNEP1 phosphatase affect interphase nuclear pore complex insertion by lipid-dependent and lipid-independent mechanisms.EMBO J. 2021 Sep 1;40(17):e106914. doi: 10.15252/embj.2020106914. Epub 2021 Jul 27. EMBO J. 2021. PMID: 34313336 Free PMC article.
-
Research Trends in C-Terminal Domain Nuclear Envelope Phosphatase 1.Life (Basel). 2023 Jun 7;13(6):1338. doi: 10.3390/life13061338. Life (Basel). 2023. PMID: 37374122 Free PMC article. Review.
-
Myosin light chain phosphatase: subunit composition, interactions and regulation.J Muscle Res Cell Motil. 1998 May;19(4):325-41. doi: 10.1023/a:1005385302064. J Muscle Res Cell Motil. 1998. PMID: 9635276 Review.
Cited by
-
Lipin phosphatidic acid phosphatases: Structure, function, regulation, and disease association.Adv Biol Regul. 2025 May;96:101082. doi: 10.1016/j.jbior.2025.101082. Epub 2025 Feb 10. Adv Biol Regul. 2025. PMID: 39948013 Review.
-
The CTR hydrophobic residues of Nem1 catalytic subunit are required to form a protein phosphatase complex with Spo7 to activate yeast Pah1 PA phosphatase.J Biol Chem. 2024 Dec;300(12):108003. doi: 10.1016/j.jbc.2024.108003. Epub 2024 Nov 17. J Biol Chem. 2024. PMID: 39551141 Free PMC article.
-
Differential reliance of CTD-nuclear envelope phosphatase 1 on its regulatory subunit in ER lipid synthesis and storage.Mol Biol Cell. 2024 Jul 1;35(7):ar101. doi: 10.1091/mbc.E23-09-0382. Epub 2024 May 22. Mol Biol Cell. 2024. PMID: 38776127 Free PMC article.
References
-
- Harris T. E., et al. , Insulin controls subcellular localization and multisite phosphorylation of the phosphatidic acid phosphatase, lipin 1. J. Biol. Chem. 282, 277–286 (2007). - PubMed
MeSH terms
Substances
Grants and funding
- T32GM722345/HHS | NIH | National Institute of General Medical Sciences (NIGMS)
- na/Alfred P. Sloan Foundation (APSF)
- R01GM131004/HHS | NIH | National Institute of General Medical Sciences (NIGMS)
- R35GM128666/HHS | NIH | National Institute of General Medical Sciences (NIGMS)
- R01 GM131004/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
