

Cellular dynamics in pig-to-human kidney xenotransplantation
- PMID: 38776915
- PMCID: PMC11317223
- DOI: 10.1016/j.medj.2024.05.003
Cellular dynamics in pig-to-human kidney xenotransplantation
Abstract
Background: Xenotransplantation of genetically engineered porcine organs has the potential to address the challenge of organ donor shortage. Two cases of porcine-to-human kidney xenotransplantation were performed, yet the physiological effects on the xenografts and the recipients' immune responses remain largely uncharacterized.
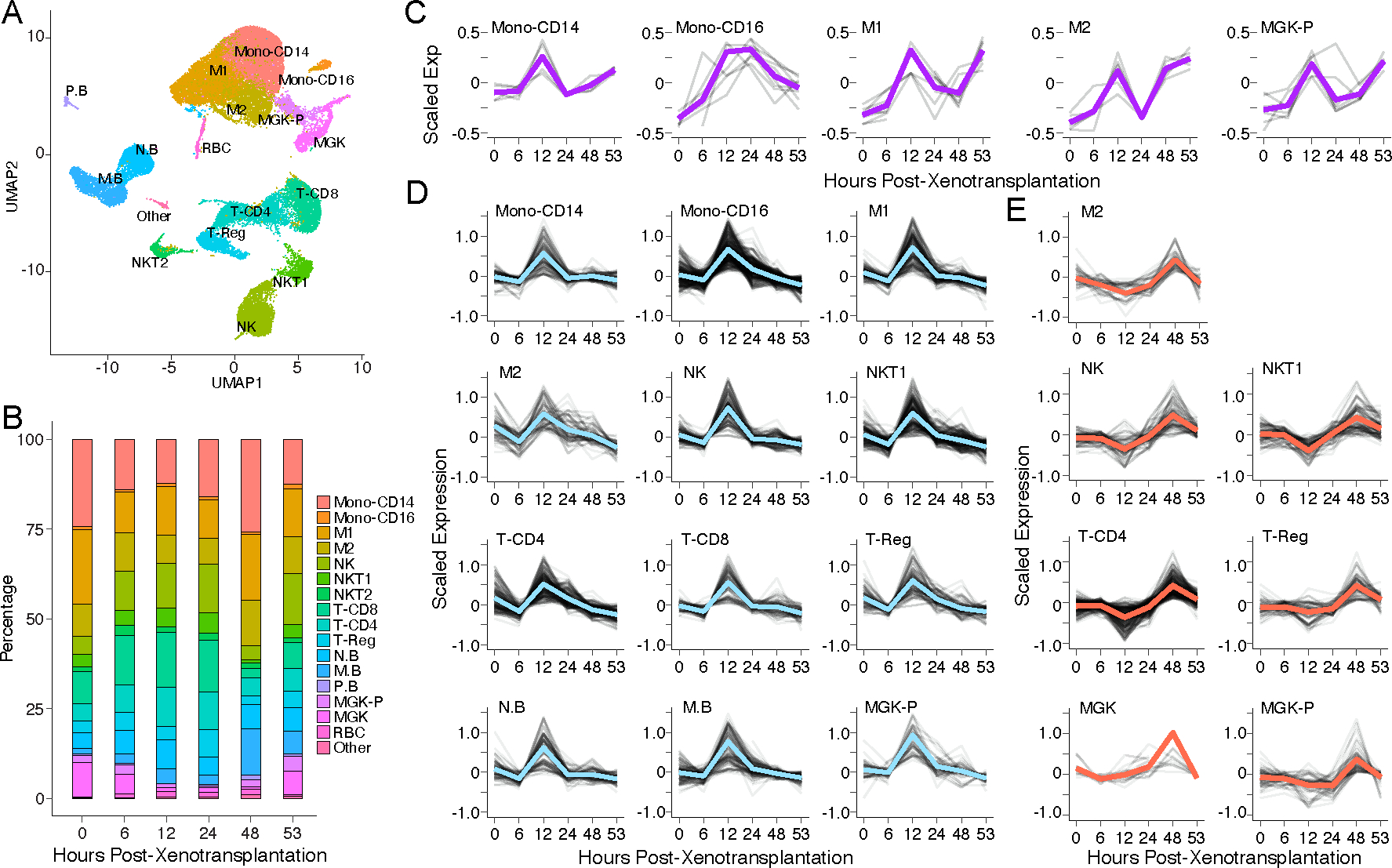
Methods: We performed single-cell RNA sequencing (scRNA-seq) and longitudinal RNA-seq analyses of the porcine kidneys to dissect xenotransplantation-associated cellular dynamics and xenograft-recipient interactions. We additionally performed longitudinal scRNA-seq of the peripheral blood mononuclear cells (PBMCs) to detect recipient immune responses across time.
Findings: Although no hyperacute rejection signals were detected, scRNA-seq analyses of the xenografts found evidence of endothelial cell and immune response activation, indicating early signs of antibody-mediated rejection. Tracing the cells' species origin, we found human immune cell infiltration in both xenografts. Human transcripts in the longitudinal bulk RNA-seq revealed that human immune cell infiltration and the activation of interferon-gamma-induced chemokine expression occurred by 12 and 48 h post-xenotransplantation, respectively. Concordantly, longitudinal scRNA-seq of PBMCs also revealed two phases of the recipients' immune responses at 12 and 48-53 h. Lastly, we observed global expression signatures of xenotransplantation-associated kidney tissue damage in the xenografts. Surprisingly, we detected a rapid increase of proliferative cells in both xenografts, indicating the activation of the porcine tissue repair program.
Conclusions: Longitudinal and single-cell transcriptomic analyses of porcine kidneys and the recipient's PBMCs revealed time-resolved cellular dynamics of xenograft-recipient interactions during xenotransplantation. These cues can be leveraged for designing gene edits and immunosuppression regimens to optimize xenotransplantation outcomes.
Funding: This work was supported by NIH RM1HG009491 and DP5OD033430.
Keywords: Translation to patients; antibody-mediated rejection; cell proliferation; genetic engineering; immune response; longitudinal RNA-seq; porcine kidney; scRNA-seq; tissue repair; xenotransplantation.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.D.B. is a founder and director of CDI Labs, Inc.; a founder of and consultant to Opentrons LabWorks/Neochromosome, Inc.; and serves or served on the scientific advisory boards of the following: CZ Biohub New York, LLC; Logomix, Inc.; Modern Meadow, Inc.; Rome Therapeutics, Inc.; Sangamo, Inc.; Tessera Therapeutics, Inc.; and the Wyss Institute. R.A.M. is on scientific advisory boards for eGenesis, Sanofi, Regeneron, CareDx, and Hansa Biopharma; is a consultant to Recombinetics; reports consulting fees from Hansa Medical, Regeneron, Thermo Fisher Scientific, Genentech, CareDx, One Lambda, ITB Med, Sanofi, and PPD Development; and reports grant support from Hansa Biopharma, all unrelated to the present work. R.A.M. also reports grant support from United Therapeutics Corporation, PBC. All other authors have no competing interests.
Figures




References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
