

The CD8+ T cell tolerance checkpoint triggers a distinct differentiation state defined by protein translation defects
- PMID: 38776918
- PMCID: PMC11807353
- DOI: 10.1016/j.immuni.2024.04.026
The CD8+ T cell tolerance checkpoint triggers a distinct differentiation state defined by protein translation defects
Abstract
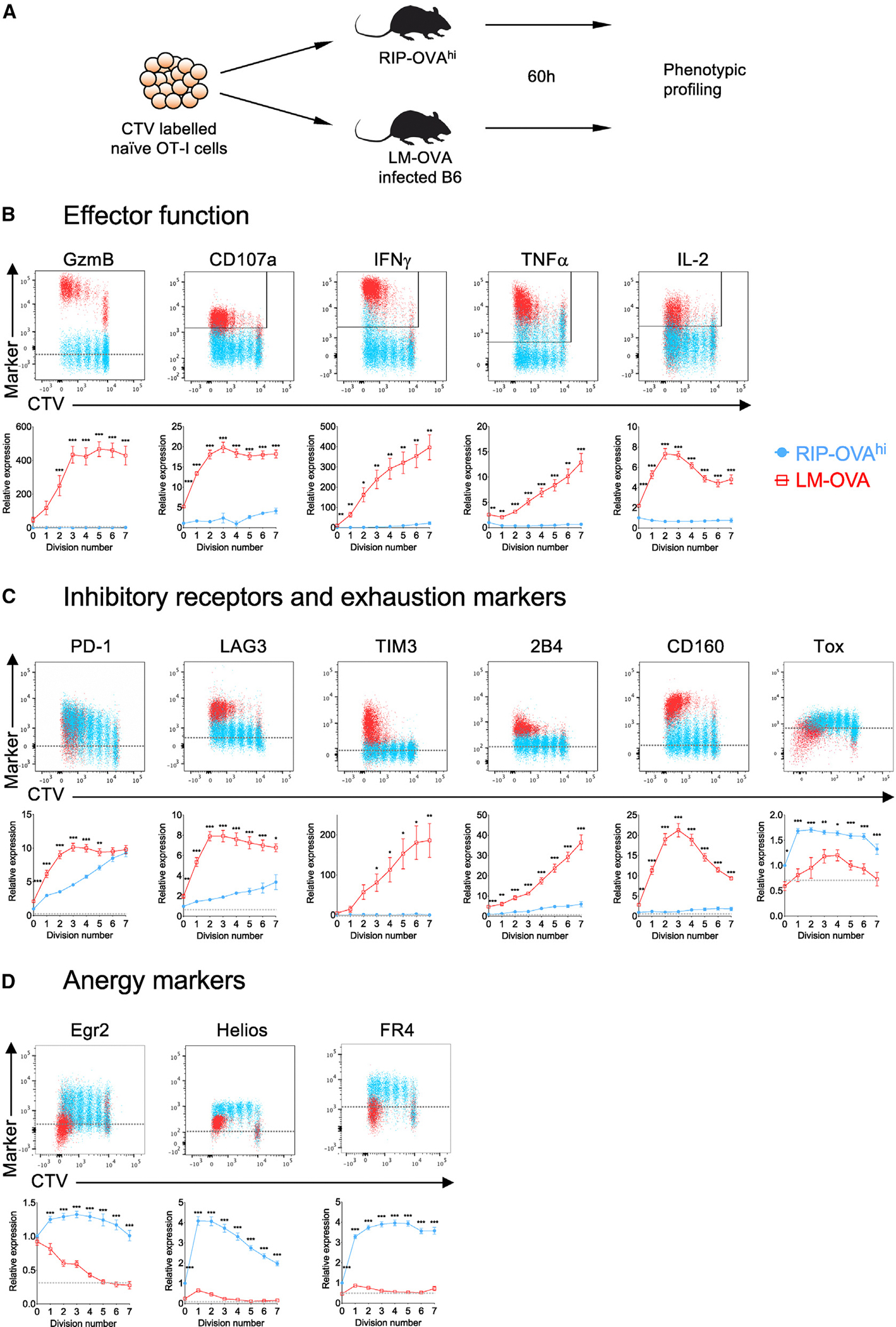
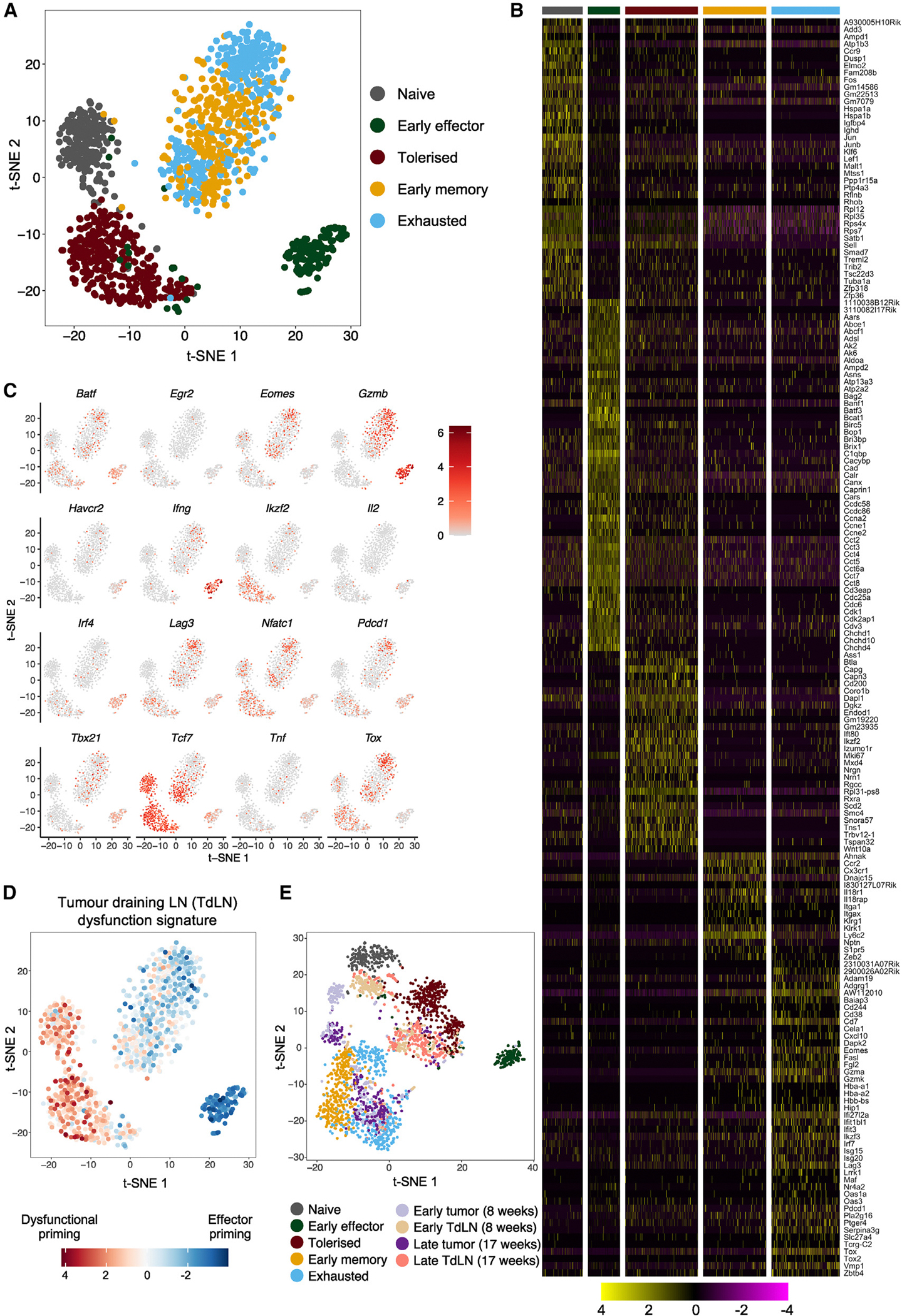
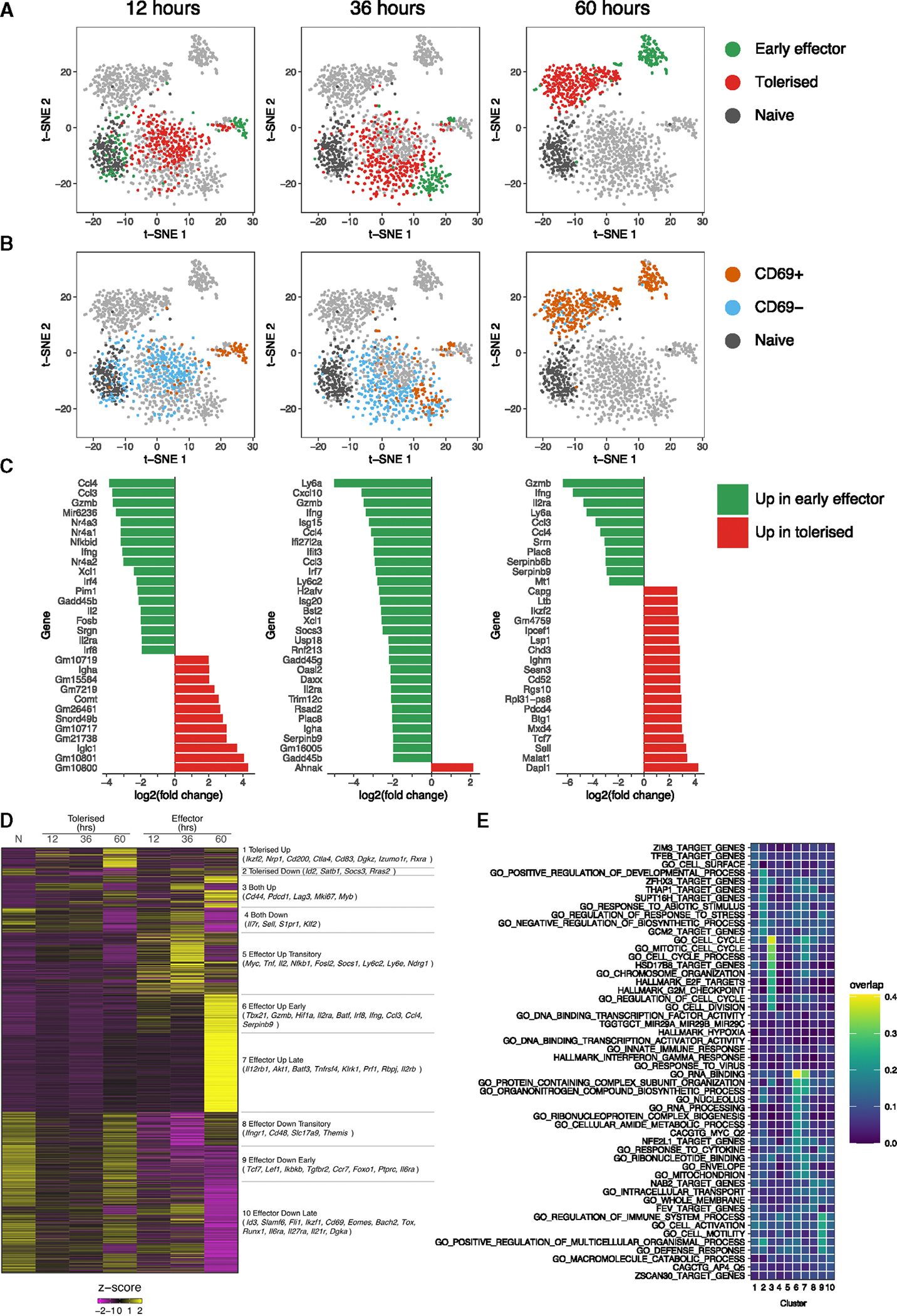
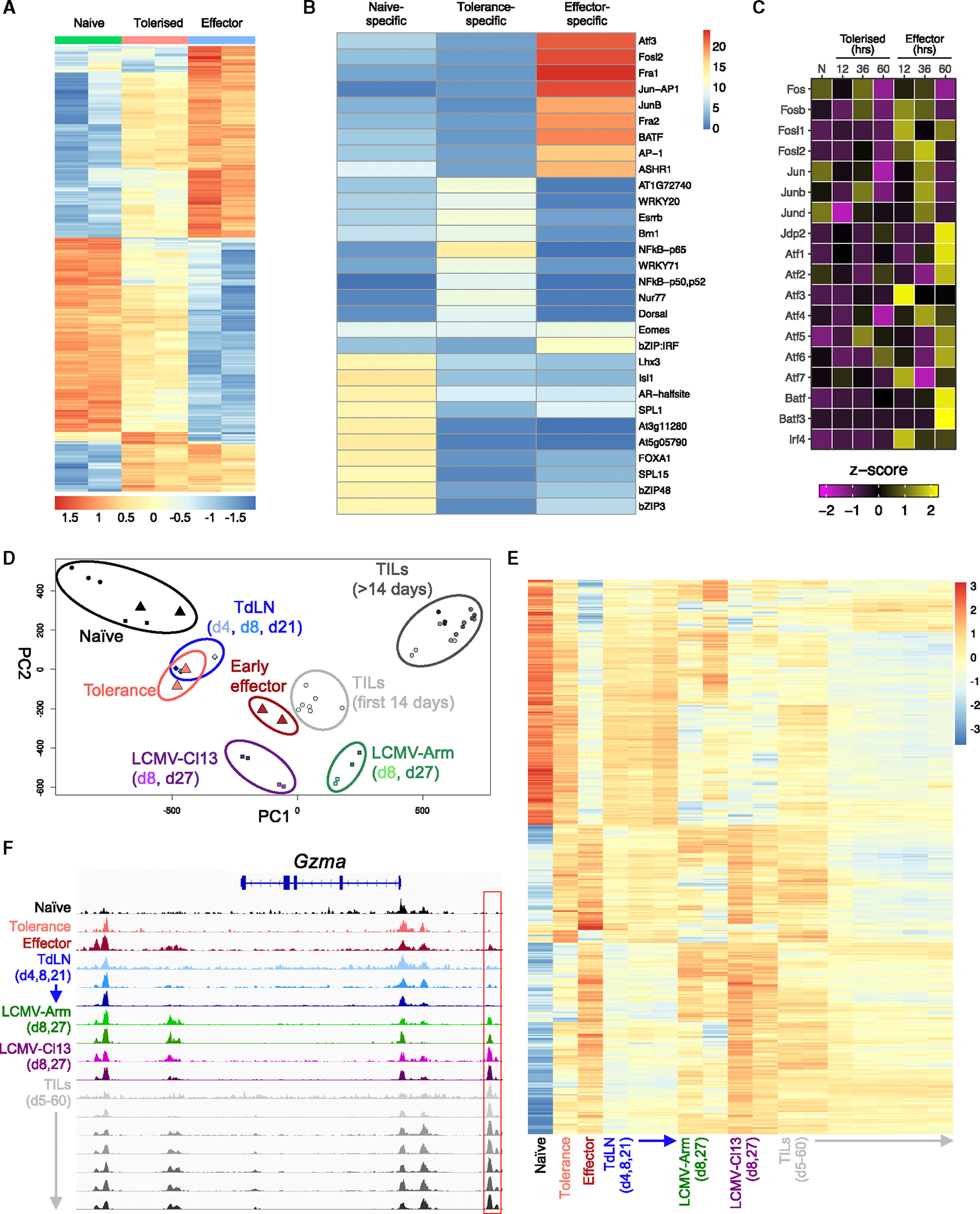
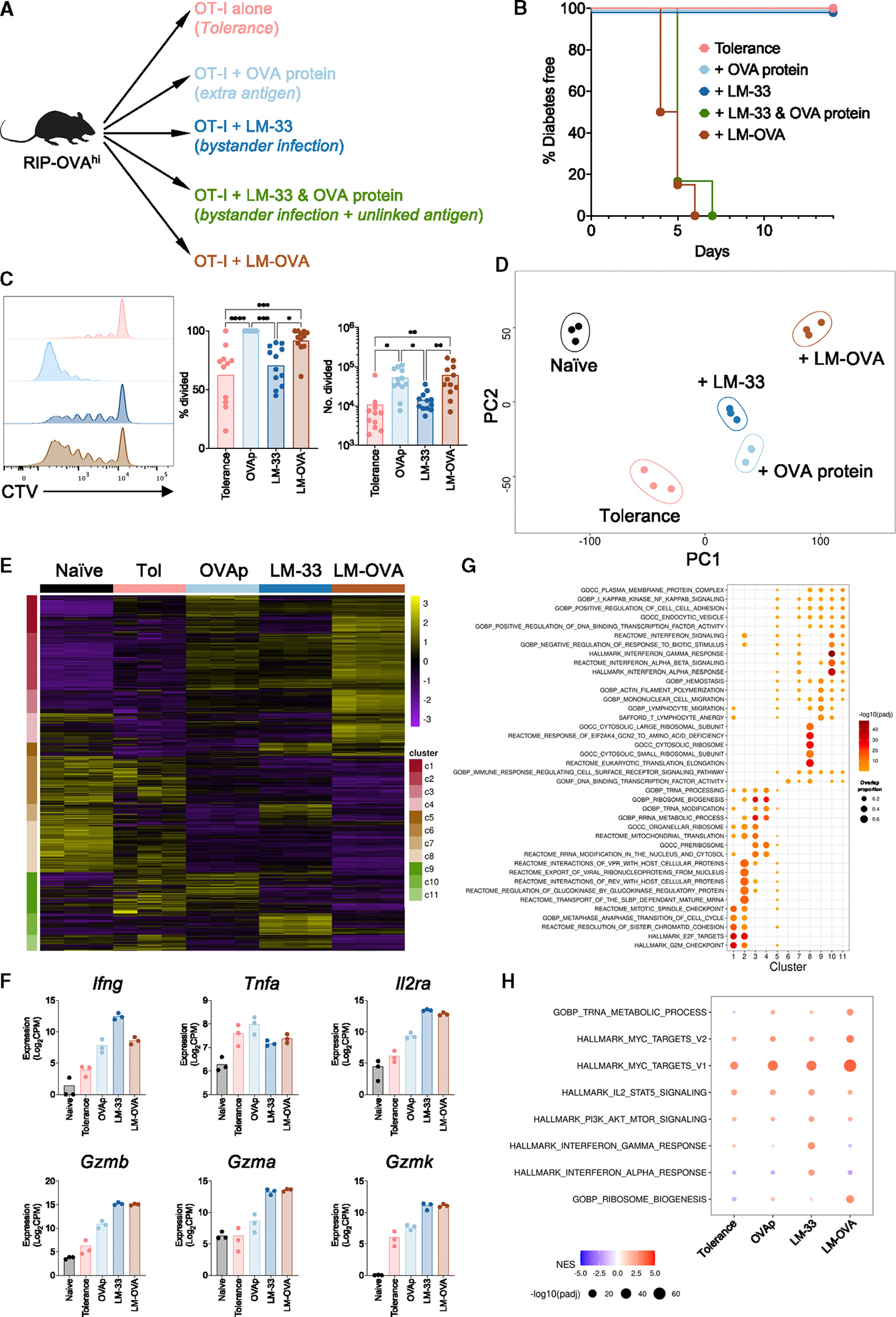
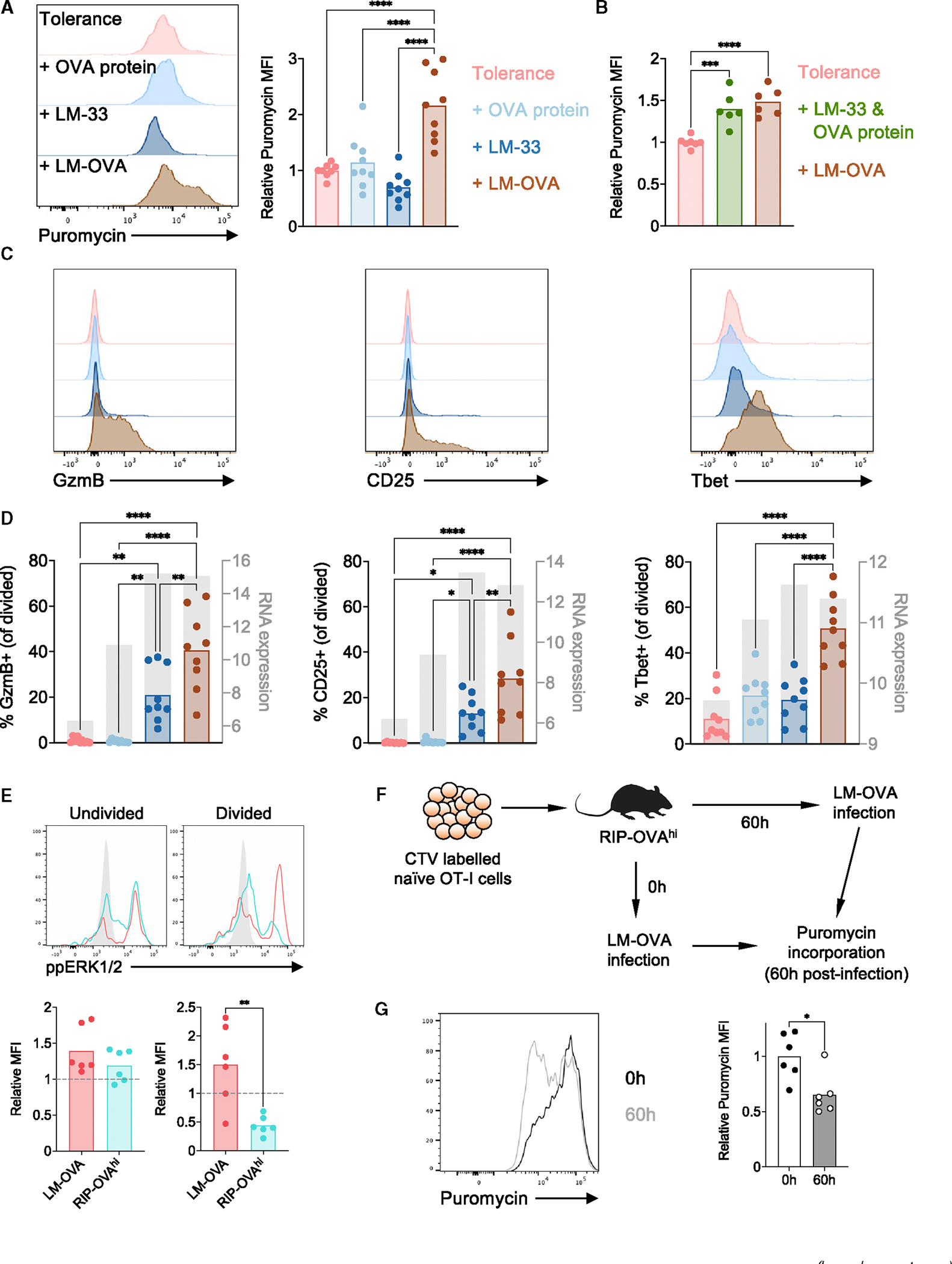
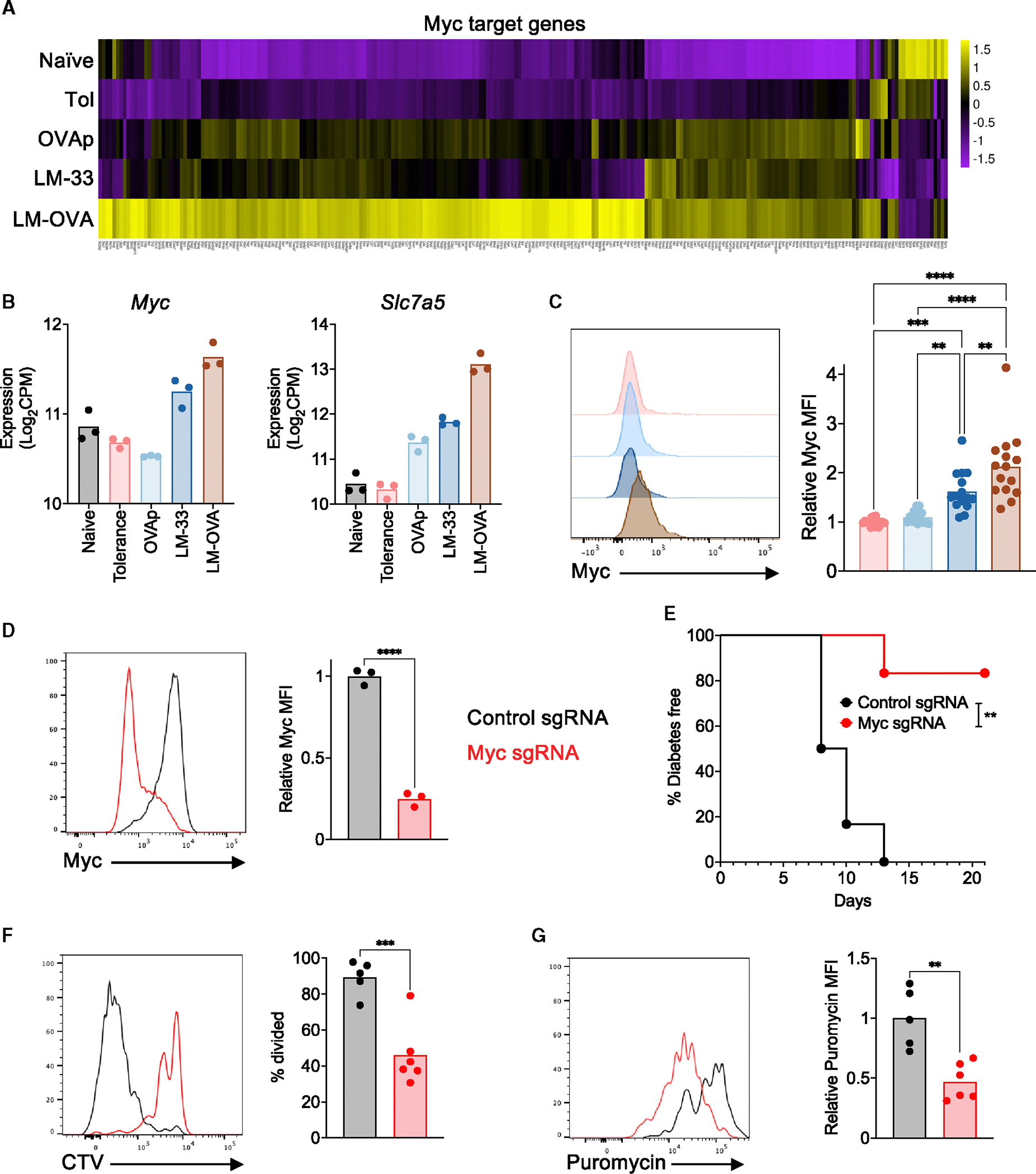
Peripheral CD8+ T cell tolerance is a checkpoint in both autoimmune disease and anti-cancer immunity. Despite its importance, the relationship between tolerance-induced states and other CD8+ T cell differentiation states remains unclear. Using flow cytometric phenotyping, single-cell RNA sequencing (scRNA-seq), and chromatin accessibility profiling, we demonstrated that in vivo peripheral tolerance to a self-antigen triggered a fundamentally distinct differentiation state separate from exhaustion, memory, and functional effector cells but analogous to cells defectively primed against tumors. Tolerant cells diverged early and progressively from effector cells, adopting a transcriptionally and epigenetically distinct state within 60 h of antigen encounter. Breaching tolerance required the synergistic actions of strong T cell receptor (TCR) signaling and inflammation, which cooperatively induced gene modules that enhanced protein translation. Weak TCR signaling during bystander infection failed to breach tolerance due to the uncoupling of effector gene expression from protein translation. Thus, tolerance engages a distinct differentiation trajectory enforced by protein translation defects.
Keywords: CD8(+) T cell; autoimmunity; cancer; differentiation; dysfunction; effector T cell; exhaustion; immunotherapy; stem-like T cell; tolerance.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests I.A.P. receives research funding from AstraZeneca and Bristol-Myers Squibb. H.Y.C. is a co-founder of Accent Therapeutics, Boundless Bio, Cartography Biosciences, and Orbital Therapeutics, and is an advisor for 10× Genomics, Arsenal Biosciences, Chroma Medicine, and Spring Discovery.
Figures
Comment in
-
CD8+ T cell tolerance: It doesn't translate.Immunity. 2024 Jun 11;57(6):1184-1186. doi: 10.1016/j.immuni.2024.05.014. Immunity. 2024. PMID: 38865962
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
