

Molecular mechanism of choline and ethanolamine transport in humans
- PMID: 38778100
- PMCID: PMC11168923
- DOI: 10.1038/s41586-024-07444-7
Molecular mechanism of choline and ethanolamine transport in humans
Abstract
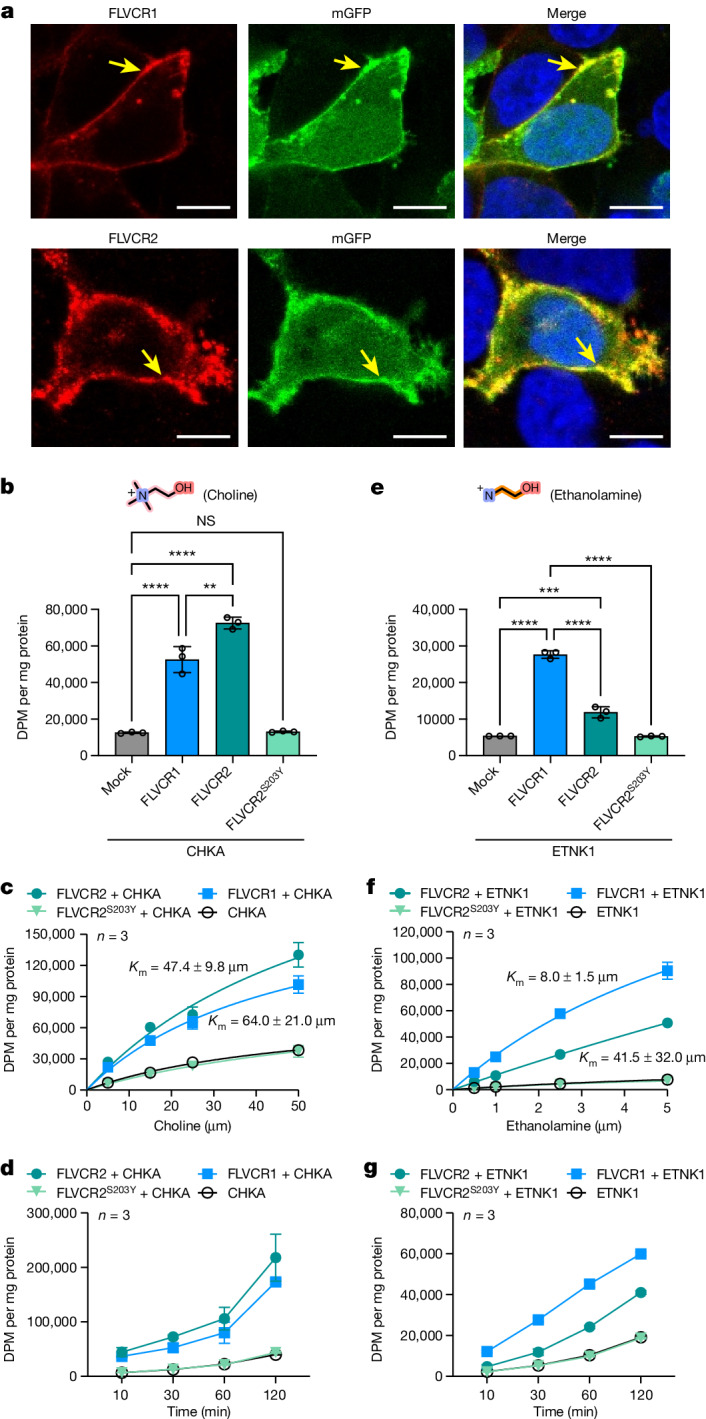
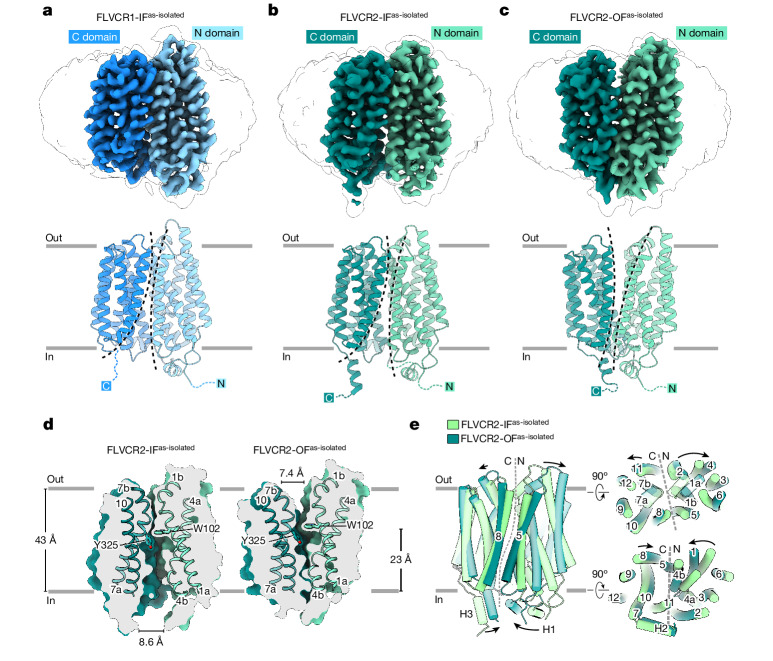
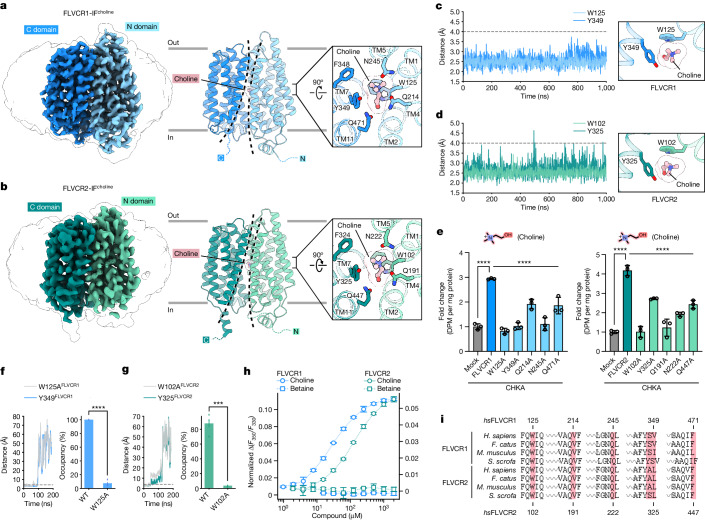
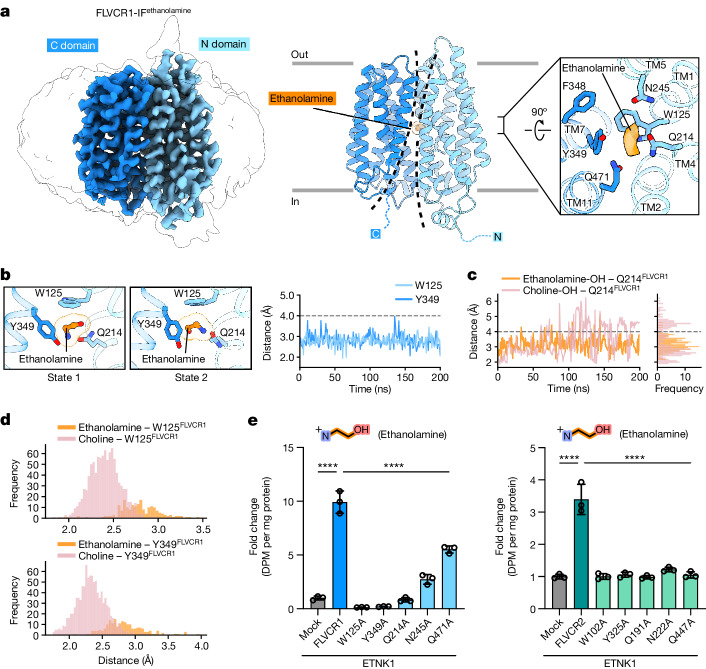
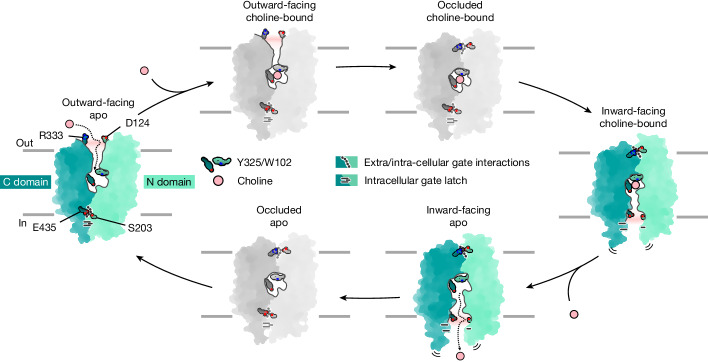
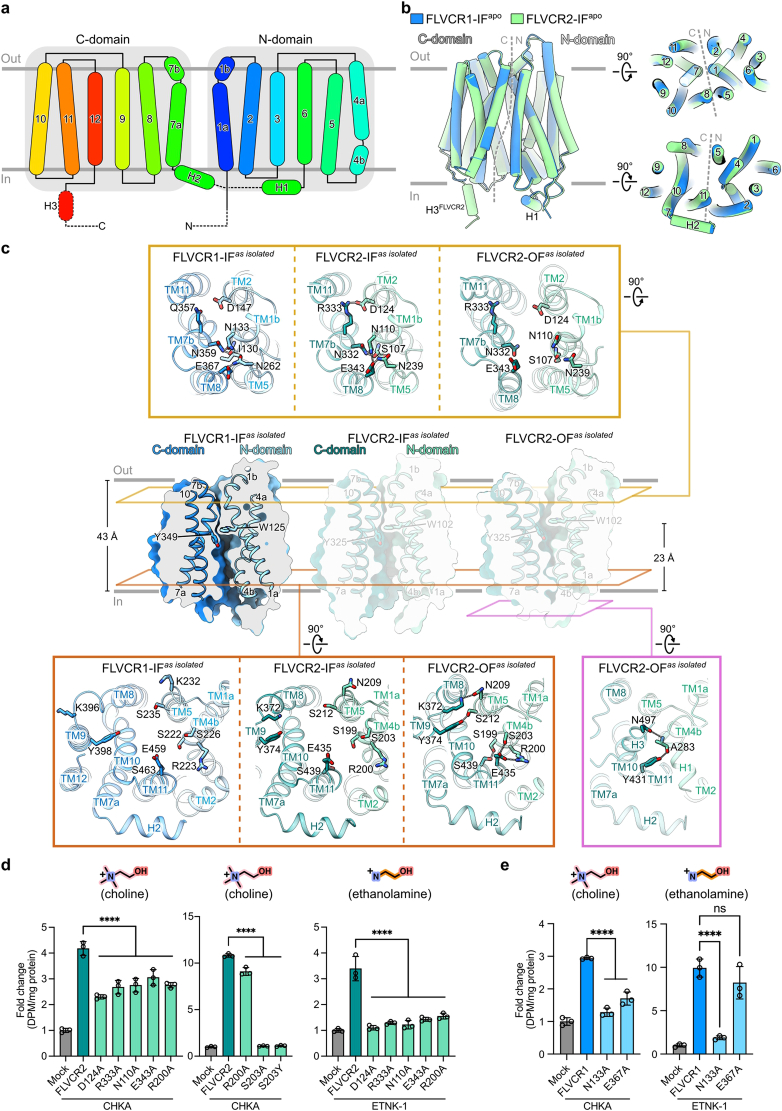
Human feline leukaemia virus subgroup C receptor-related proteins 1 and 2 (FLVCR1 and FLVCR2) are members of the major facilitator superfamily1. Their dysfunction is linked to several clinical disorders, including PCARP, HSAN and Fowler syndrome2-7. Earlier studies concluded that FLVCR1 may function as a haem exporter8-12, whereas FLVCR2 was suggested to act as a haem importer13, yet conclusive biochemical and detailed molecular evidence remained elusive for the function of both transporters14-16. Here, we show that FLVCR1 and FLVCR2 facilitate the transport of choline and ethanolamine across the plasma membrane, using a concentration-driven substrate translocation process. Through structural and computational analyses, we have identified distinct conformational states of FLVCRs and unravelled the coordination chemistry underlying their substrate interactions. Fully conserved tryptophan and tyrosine residues form the binding pocket of both transporters and confer selectivity for choline and ethanolamine through cation-π interactions. Our findings clarify the mechanisms of choline and ethanolamine transport by FLVCR1 and FLVCR2, enhance our comprehension of disease-associated mutations that interfere with these vital processes and shed light on the conformational dynamics of these major facilitator superfamily proteins during the transport cycle.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures








References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
