

Design and Semisynthesis of Biselectrophile-Functionalized Ubiquitin Probes To Investigate Transthioesterification Reactions
- PMID: 38781175
- PMCID: PMC11165569
- DOI: 10.1021/acs.orglett.4c01102
Design and Semisynthesis of Biselectrophile-Functionalized Ubiquitin Probes To Investigate Transthioesterification Reactions
Abstract
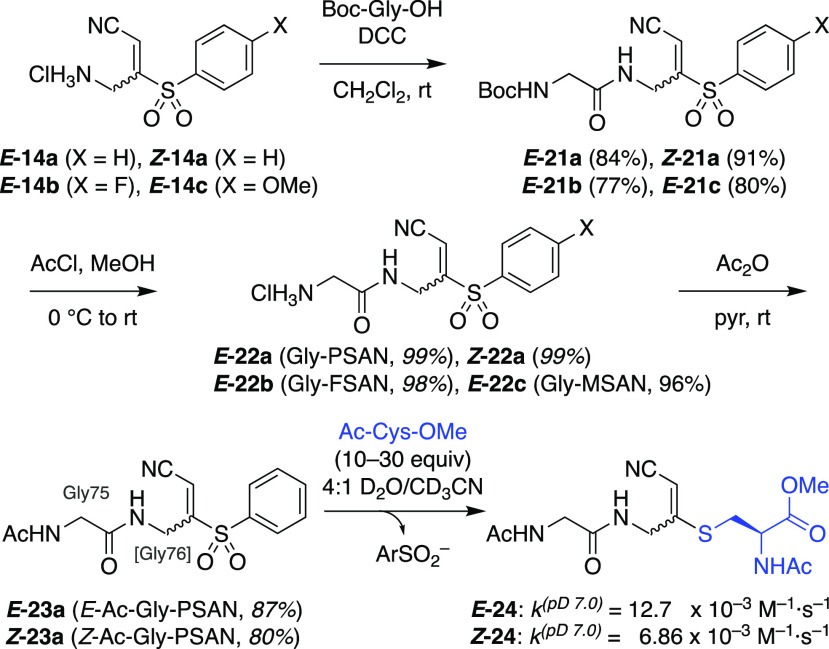
Ubiquitin (Ub) regulates a wide array of cellular processes through post-translational modification of protein substrates. Ub is conjugated at its C-terminus to target proteins via an enzymatic cascade in which covalently bound Ub thioesters are transferred from E1 activating enzymes to E2 conjugating enzymes, and then to certain E3 protein ligases. These transthioesterification reactions proceed via transient tetrahedral intermediates. A variety of chemical strategies have been used to capture E1-Ub-E2 and E2-Ub-E3 mimics, but these introduce modifications that disrupt atomic spacing at the linkage point relative to the native tetrahedral intermediate. We have developed a biselectrophilic PSAN warhead that can be installed in place of the conserved C-terminal glycine in Ub and used to form ternary protein complexes linked via cyanomethyldithioacetals that closely mimic the native tetrahedral intermediates. Investigation of the reactivity of the warhead and substituted analogues led to an effective semisynthetic route to Ub-1-PSAN, which was used to form a ternary E1-Ub*-E2 complex as a mimic of the transthioesterification intermediate.
Conflict of interest statement
The authors declare no competing financial interest.
Figures



Similar articles
-
Structural basis for transthiolation intermediates in the ubiquitin pathway.Nature. 2024 Sep;633(8028):216-223. doi: 10.1038/s41586-024-07828-9. Epub 2024 Aug 14. Nature. 2024. PMID: 39143218 Free PMC article.
-
Chemical Tools for Probing the Ub/Ubl Conjugation Cascades.Chembiochem. 2025 Jan 2;26(1):e202400659. doi: 10.1002/cbic.202400659. Epub 2024 Nov 6. Chembiochem. 2025. PMID: 39313481 Free PMC article. Review.
-
Crystal structures of an E1-E2-ubiquitin thioester mimetic reveal molecular mechanisms of transthioesterification.Nat Commun. 2021 Apr 22;12(1):2370. doi: 10.1038/s41467-021-22598-y. Nat Commun. 2021. PMID: 33888705 Free PMC article.
-
Insights into ubiquitin transfer cascades from a structure of a UbcH5B approximately ubiquitin-HECT(NEDD4L) complex.Mol Cell. 2009 Dec 25;36(6):1095-102. doi: 10.1016/j.molcel.2009.11.010. Mol Cell. 2009. PMID: 20064473 Free PMC article.
-
E2s: structurally economical and functionally replete.Biochem J. 2011 Jan 1;433(1):31-42. doi: 10.1042/BJ20100985. Biochem J. 2011. PMID: 21158740 Free PMC article. Review.
Cited by
-
Structural basis for transthiolation intermediates in the ubiquitin pathway.Nature. 2024 Sep;633(8028):216-223. doi: 10.1038/s41586-024-07828-9. Epub 2024 Aug 14. Nature. 2024. PMID: 39143218 Free PMC article.
-
Chemical Tools for Probing the Ub/Ubl Conjugation Cascades.Chembiochem. 2025 Jan 2;26(1):e202400659. doi: 10.1002/cbic.202400659. Epub 2024 Nov 6. Chembiochem. 2025. PMID: 39313481 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
