

Glial fibrillary acidic protein is pathologically modified in Alexander disease
- PMID: 38782207
- PMCID: PMC11259701
- DOI: 10.1016/j.jbc.2024.107402
Glial fibrillary acidic protein is pathologically modified in Alexander disease
Abstract
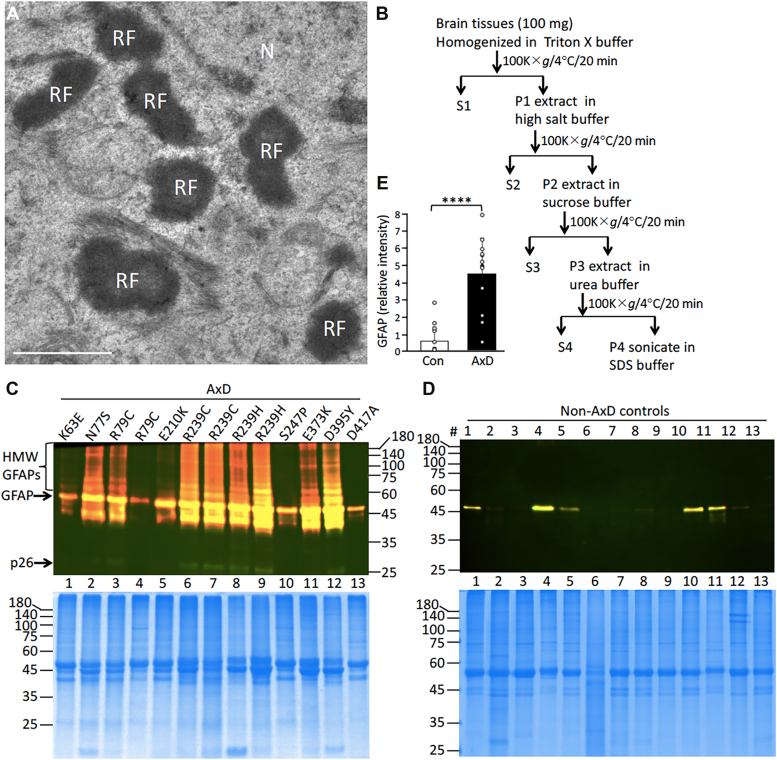
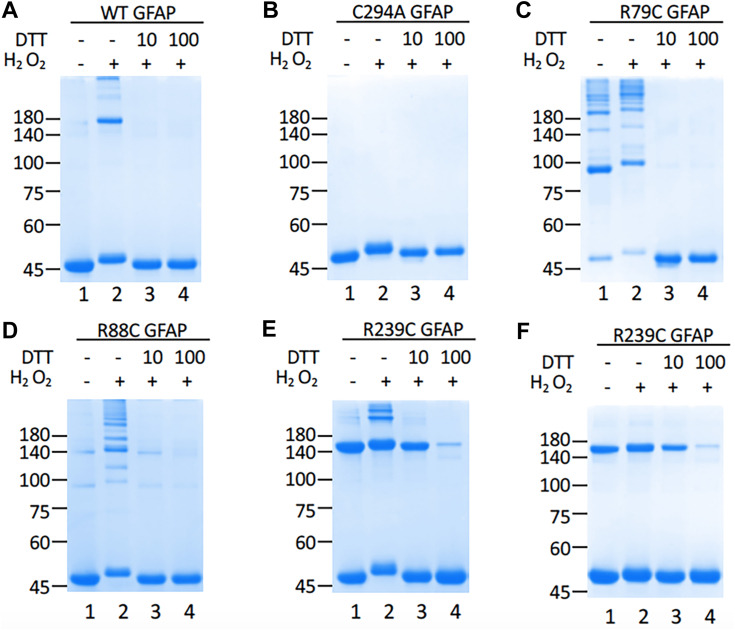
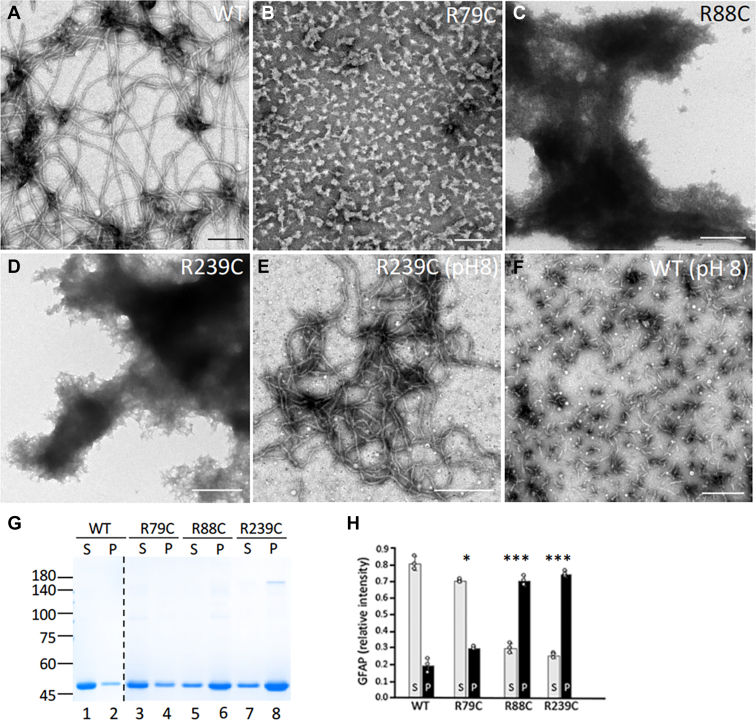
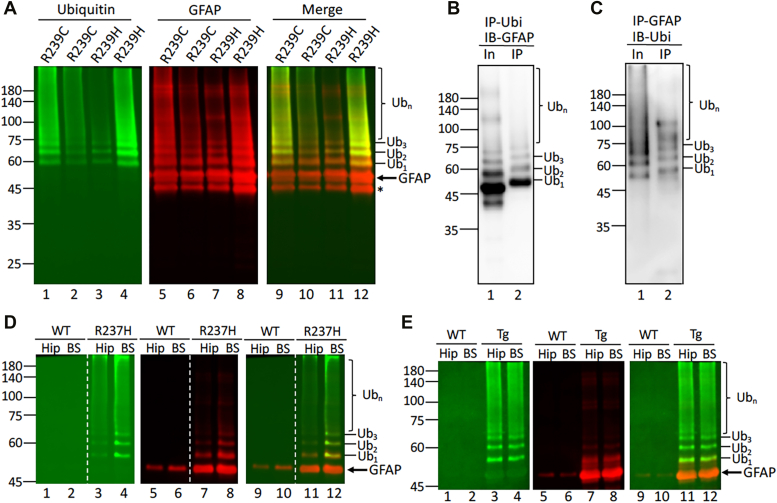
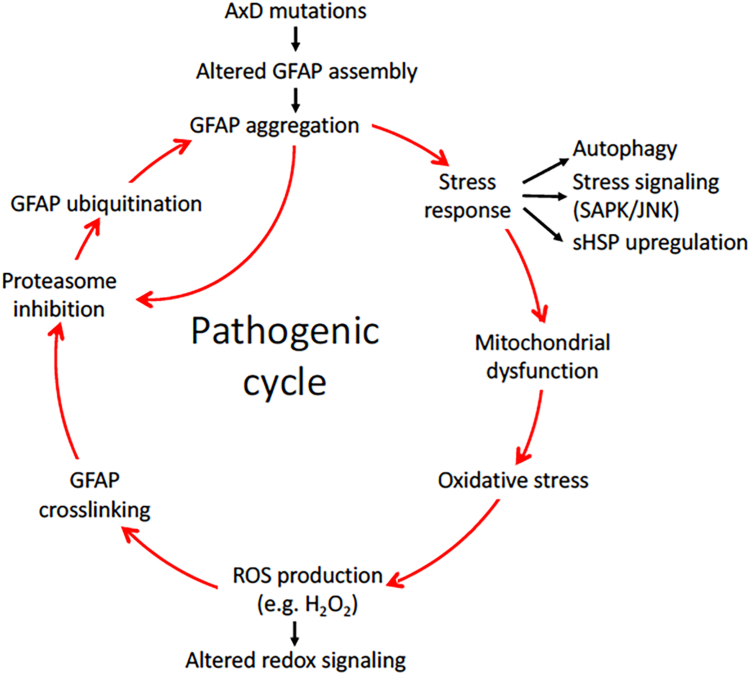
Here, we describe pathological events potentially involved in the disease pathogenesis of Alexander disease (AxD). This is a primary genetic disorder of astrocyte caused by dominant gain-of-function mutations in the gene coding for an intermediate filament protein glial fibrillary acidic protein (GFAP). Pathologically, this disease is characterized by the upregulation of GFAP and its accumulation as Rosenthal fibers. Although the genetic basis linking GFAP mutations with Alexander disease has been firmly established, the initiating events that promote GFAP accumulation and the role of Rosenthal fibers (RFs) in the disease process remain unknown. Here, we investigate the hypothesis that disease-associated mutations promote GFAP aggregation through aberrant posttranslational modifications. We found high molecular weight GFAP species in the RFs of AxD brains, indicating abnormal GFAP crosslinking as a prominent pathological feature of this disease. In vitro and cell-based studies demonstrate that cystine-generating mutations promote GFAP crosslinking by cysteine-dependent oxidation, resulting in defective GFAP assembly and decreased filament solubility. Moreover, we found GFAP was ubiquitinated in RFs of AxD patients and rodent models, supporting this modification as a critical factor linked to GFAP aggregation. Finally, we found that arginine could increase the solubility of aggregation-prone mutant GFAP by decreasing its ubiquitination and aggregation. Our study suggests a series of pathogenic events leading to AxD, involving interplay between GFAP aggregation and abnormal modifications by GFAP ubiquitination and oxidation. More important, our findings provide a basis for investigating new strategies to treat AxD by targeting abnormal GFAP modifications.
Keywords: Alexander disease; aggregation; glial fibrillary acidic protein; intermediate filament; ubiquitination.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest The authors declare that they have no conflicts of interests with the contents of this article.
Figures


Similar articles
-
The origin of Rosenthal fibers and their contributions to astrocyte pathology in Alexander disease.Acta Neuropathol Commun. 2017 Mar 31;5(1):27. doi: 10.1186/s40478-017-0425-9. Acta Neuropathol Commun. 2017. PMID: 28359321 Free PMC article.
-
Plectin regulates the organization of glial fibrillary acidic protein in Alexander disease.Am J Pathol. 2006 Mar;168(3):888-97. doi: 10.2353/ajpath.2006.051028. Am J Pathol. 2006. PMID: 16507904 Free PMC article.
-
Synemin is expressed in reactive astrocytes and Rosenthal fibers in Alexander disease.APMIS. 2014 Jan;122(1):76-80. doi: 10.1111/apm.12088. Epub 2013 Apr 18. APMIS. 2014. PMID: 23594359
-
GFAP mutations in Alexander disease.Int J Dev Neurosci. 2002 Jun-Aug;20(3-5):259-68. doi: 10.1016/s0736-5748(02)00019-9. Int J Dev Neurosci. 2002. PMID: 12175861 Review.
-
Clinical aspects and pathology of Alexander disease, and morphological and functional alteration of astrocytes induced by GFAP mutation.Neuropathology. 2012 Aug;32(4):440-6. doi: 10.1111/j.1440-1789.2011.01268.x. Epub 2011 Nov 28. Neuropathology. 2012. PMID: 22118268 Review.
Cited by
-
Unveiling the molecular blueprint of SKP-SCs-mediated tissue engineering-enhanced neuroregeneration.J Nanobiotechnology. 2024 Dec 26;22(1):796. doi: 10.1186/s12951-024-03076-1. J Nanobiotechnology. 2024. PMID: 39725969 Free PMC article.
-
"Frog face and strangulated medulla": neuroimaging phenotype in a novel mutation in GFAP gene causing adult onset Alexander disease.Neurol Sci. 2025 Aug;46(8):4065-4068. doi: 10.1007/s10072-025-08161-3. Epub 2025 Apr 8. Neurol Sci. 2025. PMID: 40198472 No abstract available.
-
Deletions in Glial Fibrillary Acidic Protein Leading to Alterations in Intermediate Filament Assembly and Network Formation.Int J Mol Sci. 2025 Feb 23;26(5):1913. doi: 10.3390/ijms26051913. Int J Mol Sci. 2025. PMID: 40076540 Free PMC article.
-
The GFAP proteoform puzzle: How to advance GFAP as a fluid biomarker in neurological diseases.J Neurochem. 2025 Jan;169(1):e16226. doi: 10.1111/jnc.16226. Epub 2024 Sep 17. J Neurochem. 2025. PMID: 39289040 Free PMC article. Review.
-
Genotype-Phenotype Association for 14 GFAP Variants in Alexander Disease.Neurol Genet. 2025 Jun 5;11(3):e200270. doi: 10.1212/NXG.0000000000200270. eCollection 2025 Jun. Neurol Genet. 2025. PMID: 40520824 Free PMC article.
References
-
- Eliasson C., Sahlgren C., Berthold C.H., Stakeberg J., Celis J.E., Betsholtz C., et al. Intermediate filament protein partnership in astrocytes. J. Biol. Chem. 1999;274:23996–24006. - PubMed
-
- Thomsen R., Pallesen J., Daugaard T.F., Borglum A.D., Nielsen A.L. Genome wide assessment of mRNA in astrocyte protrusions by direct RNA sequencing reveals mRNA localization for the intermediate filament protein nestin. Glia. 2013;61:1922–1937. - PubMed
-
- Jing R., Wilhelmsson U., Goodwill W., Li L., Pan Y., Pekny M., et al. Synemin is expressed in reactive astrocytes in neurotrauma and interacts differentially with vimentin and GFAP intermediate filament networks. J. Cell Sci. 2007;120:1267–1277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
