

Angle between DNA linker and nucleosome core particle regulates array compaction revealed by individual-particle cryo-electron tomography
- PMID: 38782894
- PMCID: PMC11116431
- DOI: 10.1038/s41467-024-48305-1
Angle between DNA linker and nucleosome core particle regulates array compaction revealed by individual-particle cryo-electron tomography
Abstract
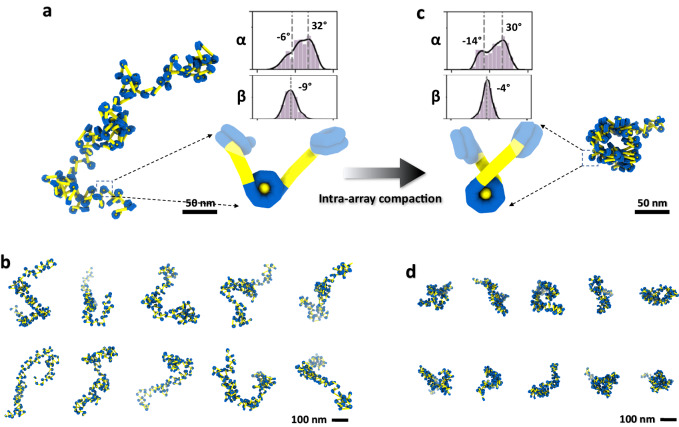
The conformational dynamics of nucleosome arrays generate a diverse spectrum of microscopic states, posing challenges to their structural determination. Leveraging cryogenic electron tomography (cryo-ET), we determine the three-dimensional (3D) structures of individual mononucleosomes and arrays comprising di-, tri-, and tetranucleosomes. By slowing the rate of condensation through a reduction in ionic strength, we probe the intra-array structural transitions that precede inter-array interactions and liquid droplet formation. Under these conditions, the arrays exhibite irregular zig-zag conformations with loose packing. Increasing the ionic strength promoted intra-array compaction, yet we do not observe the previously reported regular 30-nanometer fibers. Interestingly, the presence of H1 do not induce array compaction; instead, one-third of the arrays display nucleosomes invaded by foreign DNA, suggesting an alternative role for H1 in chromatin network construction. We also find that the crucial parameter determining the structure adopted by chromatin arrays is the angle between the entry and exit of the DNA and the corresponding tangents to the nucleosomal disc. Our results provide insights into the initial stages of intra-array compaction, a critical precursor to condensation in the regulation of chromatin organization.
© 2024. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
The authors declare no competing interests.
Figures




Update of
-
Conformational Change of Nucleosome Arrays prior to Phase Separation.Res Sq [Preprint]. 2023 Jan 19:rs.3.rs-2460504. doi: 10.21203/rs.3.rs-2460504/v1. Res Sq. 2023. Update in: Nat Commun. 2024 May 23;15(1):4395. doi: 10.1038/s41467-024-48305-1. PMID: 36711774 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
- DE-SC0022305/DOE | SC | Basic Energy Sciences (BES)
- R01GM104427/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- DE-AC02-05CH11231/DOE | SC | Basic Energy Sciences (BES)
- DE-AC02-05CH11231/DOE | Office of Science (SC)
- R01 DK042667/DK/NIDDK NIH HHS/United States
- R01MH077303/U.S. Department of Health & Human Services | NIH | National Institute of Mental Health (NIMH)
- R01GM032543/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R01 GM104427/GM/NIGMS NIH HHS/United States
- R01HL115153/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- R01 HL115153/HL/NHLBI NIH HHS/United States
- R01 GM032543/GM/NIGMS NIH HHS/United States
- R37 GM032543/GM/NIGMS NIH HHS/United States
- R01 MH077303/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
