

Disrupting Na+ ion homeostasis and Na+/K+ ATPase activity in breast cancer cells directly modulates glycolysis in vitro and in vivo
- PMID: 38783368
- PMCID: PMC11119389
- DOI: 10.1186/s40170-024-00343-5
Disrupting Na+ ion homeostasis and Na+/K+ ATPase activity in breast cancer cells directly modulates glycolysis in vitro and in vivo
Abstract
Background: Glycolytic flux is regulated by the energy demands of the cell. Upregulated glycolysis in cancer cells may therefore result from increased demand for adenosine triphosphate (ATP), however it is unknown what this extra ATP turnover is used for. We hypothesise that an important contribution to the increased glycolytic flux in cancer cells results from the ATP demand of Na+/K+-ATPase (NKA) due to altered sodium ion homeostasis in cancer cells.
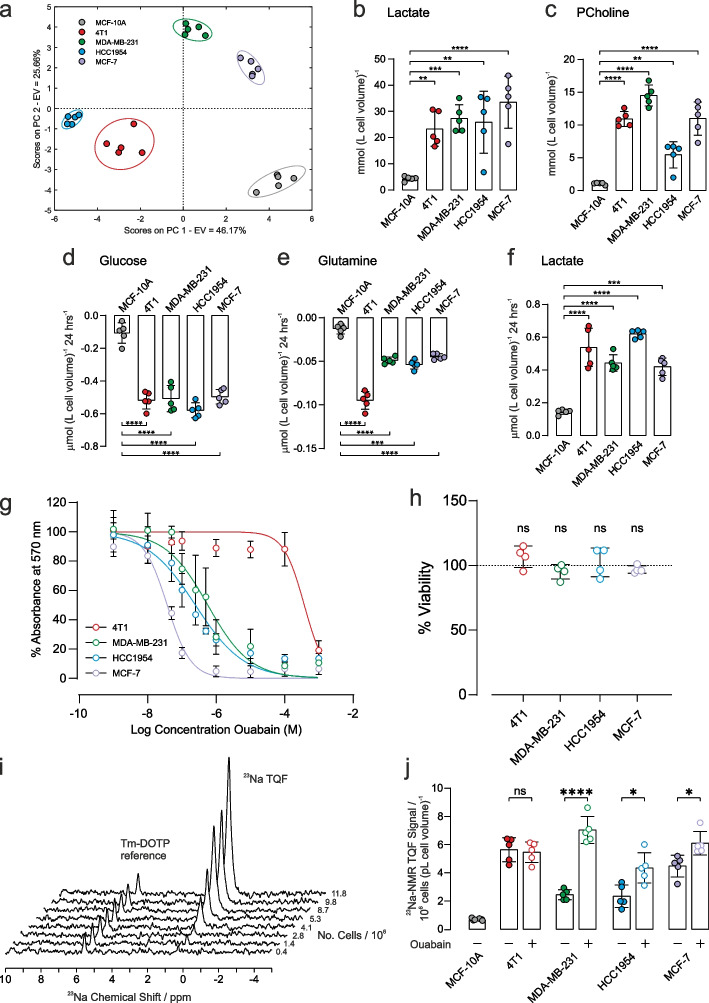
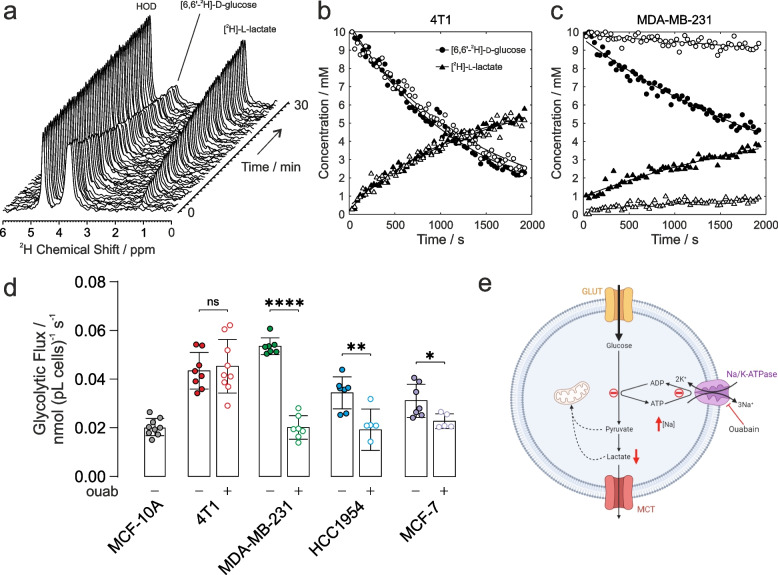
Methods: Live whole-cell measurements of intracellular sodium [Na+]i were performed in three human breast cancer cells (MDA-MB-231, HCC1954, MCF-7), in murine breast cancer cells (4T1), and control human epithelial cells MCF-10A using triple quantum filtered 23Na nuclear magnetic resonance (NMR) spectroscopy. Glycolytic flux was measured by 2H NMR to monitor conversion of [6,6-2H2]D-glucose to [2H]-labelled L-lactate at baseline and in response to NKA inhibition with ouabain. Intracellular [Na+]i was titrated using isotonic buffers with varying [Na+] and [K+] and introducing an artificial Na+ plasma membrane leak using the ionophore gramicidin-A. Experiments were carried out in parallel with cell viability assays, 1H NMR metabolomics of intracellular and extracellular metabolites, extracellular flux analyses and in vivo measurements in a MDA-MB-231 human-xenograft mouse model using 2-deoxy-2-[18F]fluoroglucose (18F-FDG) positron emission tomography (PET).
Results: Intracellular [Na+]i was elevated in human and murine breast cancer cells compared to control MCF-10A cells. Acute inhibition of NKA by ouabain resulted in elevated [Na+]i and inhibition of glycolytic flux in all three human cancer cells which are ouabain sensitive, but not in the murine cells which are ouabain resistant. Permeabilization of cell membranes with gramicidin-A led to a titratable increase of [Na+]i in MDA-MB-231 and 4T1 cells and a Na+-dependent increase in glycolytic flux. This was attenuated with ouabain in the human cells but not in the murine cells. 18FDG PET imaging in an MDA-MB-231 human-xenograft mouse model recorded lower 18FDG tumour uptake when treated with ouabain while murine tissue uptake was unaffected.
Conclusions: Glycolytic flux correlates with Na+-driven NKA activity in breast cancer cells, providing evidence for the 'centrality of the [Na+]i-NKA nexus' in the mechanistic basis of the Warburg effect.
Keywords: Breast cancer; Glycolysis; Intracellular sodium; NaK ATPase; Ouabain; Warburg effect.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Warburg O. Über den Stoffwechsel der Carcinomzelle. Naturwissenschaften. 1924;12(50):1131–1137. doi: 10.1007/BF01504608. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
