

Stress-impaired reward pathway promotes distinct feeding behavior patterns
- PMID: 38784098
- PMCID: PMC11111882
- DOI: 10.3389/fnins.2024.1349366
Stress-impaired reward pathway promotes distinct feeding behavior patterns
Abstract
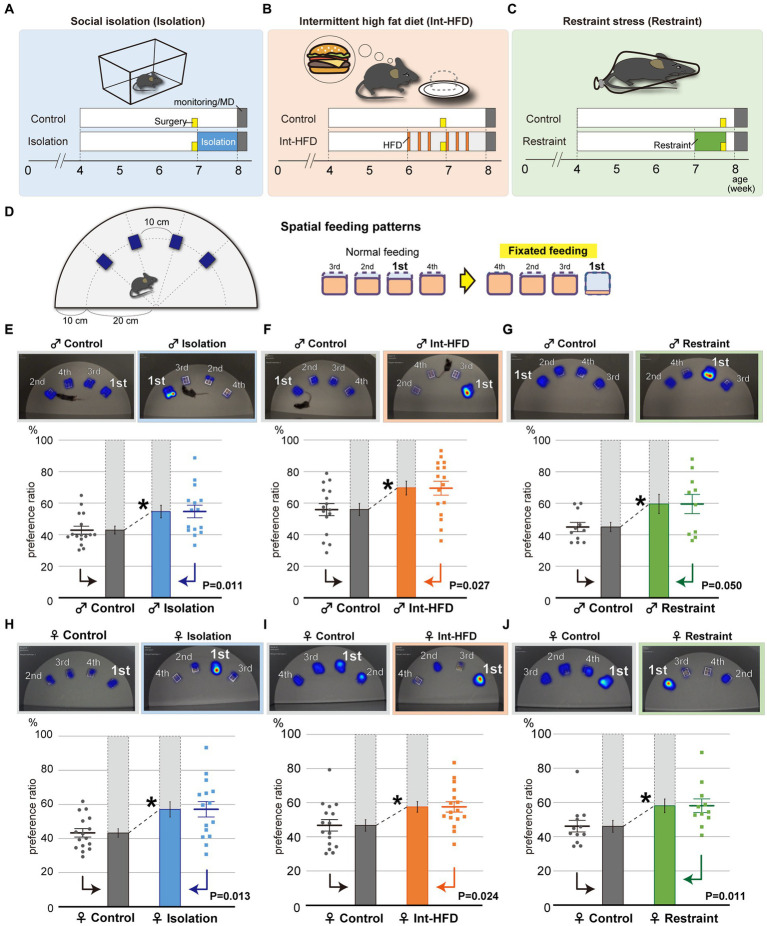
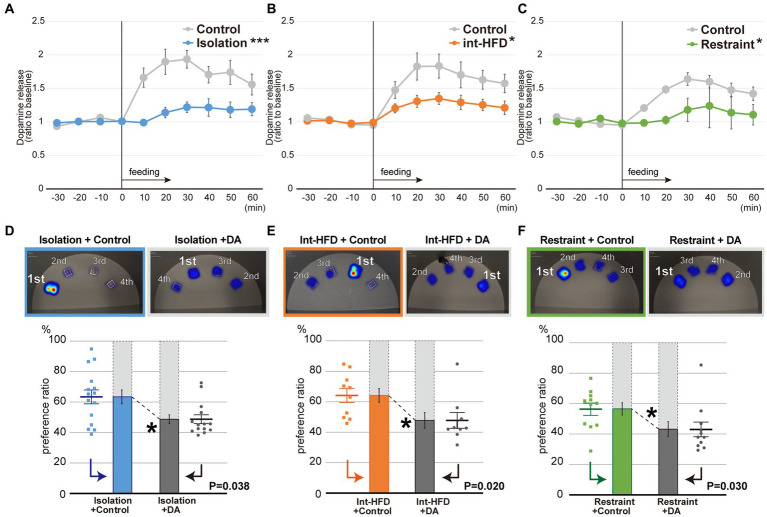
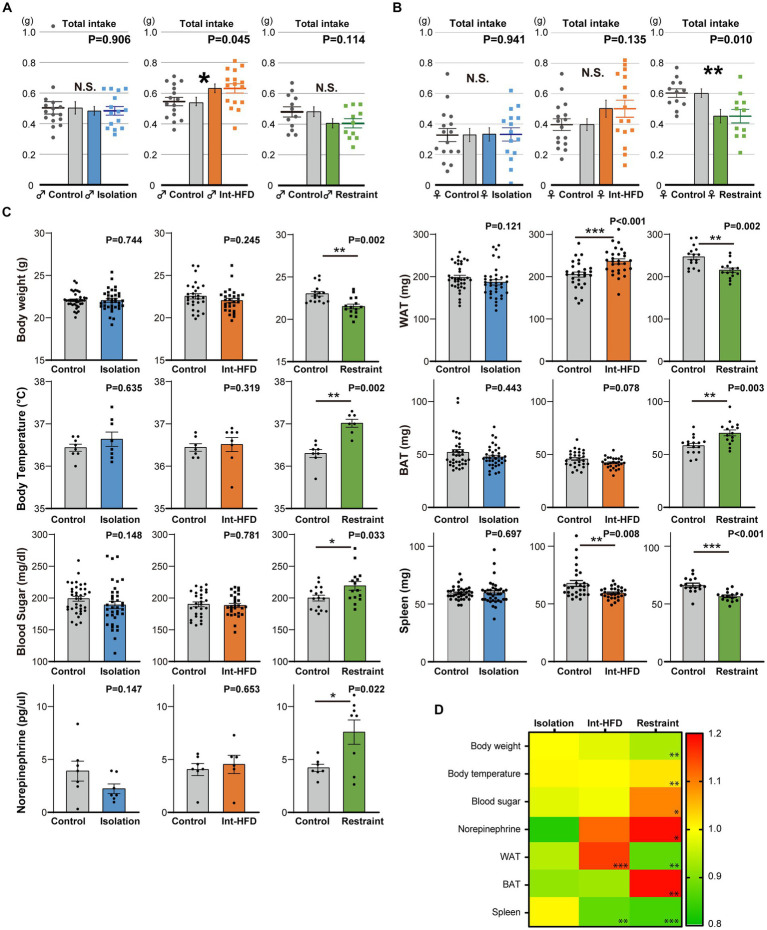
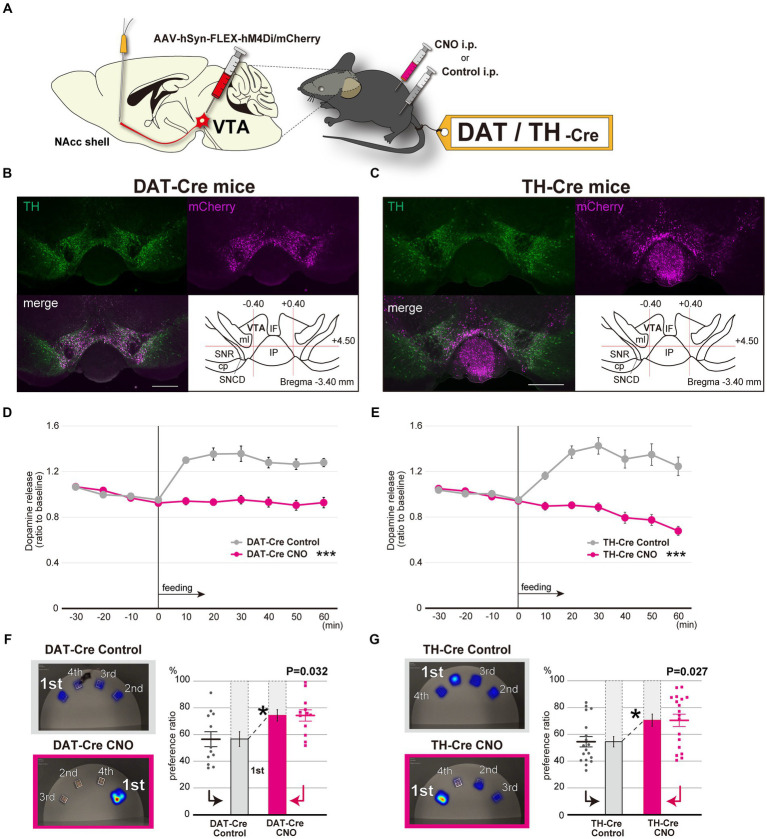
Although dietary behaviors are affected by neuropsychiatric disorders, various environmental conditions can have strong effects as well. We found that mice under multiple stresses, including social isolation, intermittent high-fat diet, and physical restraint, developed feeding behavior patterns characterized by a deviated bait approach (fixated feeding). All the tested stressors affected dopamine release at the nucleus accumbens (NAcc) shell and dopamine normalization reversed the feeding defects. Moreover, inhibition of dopaminergic activity in the ventral tegmental area that projects into the NAcc shell caused similar feeding pattern aberrations. Given that the deviations were not consistently accompanied by changes in the amount consumed or metabolic factors, the alterations in feeding behaviors likely reflect perturbations to a critical stress-associated pathway in the mesolimbic dopamine system. Thus, deviations in feeding behavior patterns that reflect reward system abnormalities can be sensitive biomarkers of psychosocial and physical stress.
Keywords: biomarker; dopamine; feeding behavior patterns; fixated feeding; psychosocial stress; reward system.
Copyright © 2024 Fujioka, Kawai, Endo, Ishibashi, Iwade, Tuerde, Kaibuchi, Yamashita, Yamanaka, Katsuno, Watanabe, Sobue and Ishigaki.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Optogenetically-induced tonic dopamine release from VTA-nucleus accumbens projections inhibits reward consummatory behaviors.Neuroscience. 2016 Oct 1;333:54-64. doi: 10.1016/j.neuroscience.2016.07.006. Epub 2016 Jul 13. Neuroscience. 2016. PMID: 27421228 Free PMC article.
-
Ventral Tegmental Area Dopamine Cell Activation during Male Rat Sexual Behavior Regulates Neuroplasticity and d-Amphetamine Cross-Sensitization following Sex Abstinence.J Neurosci. 2016 Sep 21;36(38):9949-61. doi: 10.1523/JNEUROSCI.0937-16.2016. J Neurosci. 2016. PMID: 27656032 Free PMC article.
-
Transient receptor potential vanilloid 3 (TRPV3) in the ventral tegmental area of rat: Role in modulation of the mesolimbic-dopamine reward pathway.Neuropharmacology. 2016 Nov;110(Pt A):198-210. doi: 10.1016/j.neuropharm.2016.04.012. Epub 2016 Apr 12. Neuropharmacology. 2016. PMID: 27084697
-
Neuropeptides Modulate Feeding via the Dopamine Reward Pathway.Neurochem Res. 2023 Sep;48(9):2622-2643. doi: 10.1007/s11064-023-03954-4. Epub 2023 May 26. Neurochem Res. 2023. PMID: 37233918 Review.
-
Role of ghrelin in drug abuse and reward-relevant behaviors: a burgeoning field and gaps in the literature.Curr Drug Abuse Rev. 2013 Sep;6(3):231-44. doi: 10.2174/1874473707666140205200532. Curr Drug Abuse Rev. 2013. PMID: 24502454 Review.
Cited by
-
Correcting tau isoform ratios with a long-acting antisense oligonucleotide alleviates 4R-tauopathy phenotypes.Mol Ther Nucleic Acids. 2025 Mar 5;36(2):102503. doi: 10.1016/j.omtn.2025.102503. eCollection 2025 Jun 10. Mol Ther Nucleic Acids. 2025. PMID: 40206658 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases