

Mutational Analysis of RIP Type I Dianthin-30 Suggests a Role for Arg24 in Endocytosis
- PMID: 38787071
- PMCID: PMC11125672
- DOI: 10.3390/toxins16050219
Mutational Analysis of RIP Type I Dianthin-30 Suggests a Role for Arg24 in Endocytosis
Abstract
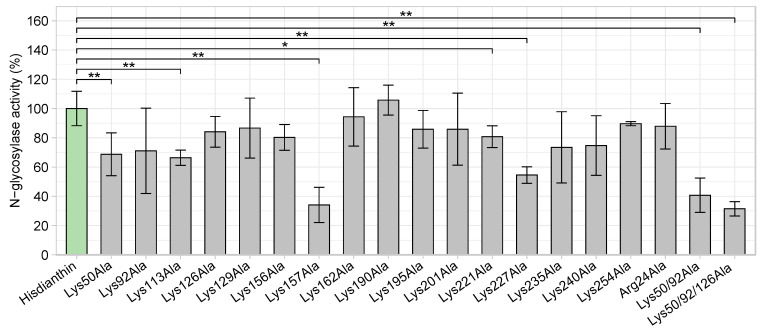
Saponin-mediated endosomal escape is a mechanism that increases the cytotoxicity of type I ribosome-inactivating proteins (type I RIPs). In order to actualize their cytotoxicity, type I RIPs must be released into the cytosol after endocytosis. Without release from the endosomes, type I RIPs are largely degraded and cannot exert their cytotoxic effects. Certain triterpene saponins are able to induce the endosomal escape of these type I RIPs, thus increasing their cytotoxicity. However, the molecular mechanism underlying the endosomal escape enhancement of type I RIPs by triterpene saponins has not been fully elucidated. In this report, we investigate the involvement of the basic amino acid residues of dianthin-30, a type I RIP isolated from the plant Dianthus caryophyllus L., in endosomal escape enhancement using alanine scanning. Therefore, we designed 19 alanine mutants of dianthin-30. Each mutant was combined with SO1861, a triterpene saponin isolated from the roots of Saponaria officinalis L., and subjected to a cytotoxicity screening in Neuro-2A cells. Cytotoxic screening revealed that dianthin-30 mutants with lysine substitutions did not impair the endosomal escape enhancement. There was one particular mutant dianthin, Arg24Ala, that exhibited significantly reduced synergistic cytotoxicity in three mammalian cell lines. However, this reduction was not based on an altered interaction with SO1861. It was, rather, due to the impaired endocytosis of dianthin Arg24Ala into the cells.
Keywords: N-glycosylase; basic amino acid residues; endocytosis; endosomal escape enhancer; ribosome-inactivating proteins; triterpene saponins; type I RIP.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures








Similar articles
-
Dianthin and Its Potential in Targeted Tumor Therapies.Toxins (Basel). 2019 Oct 11;11(10):592. doi: 10.3390/toxins11100592. Toxins (Basel). 2019. PMID: 31614697 Free PMC article. Review.
-
Combinatorial approach to increase efficacy of Cetuximab, Panitumumab and Trastuzumab by dianthin conjugation and co-application of SO1861.Biochem Pharmacol. 2015 Oct 1;97(3):247-55. doi: 10.1016/j.bcp.2015.07.040. Epub 2015 Aug 5. Biochem Pharmacol. 2015. PMID: 26253687
-
Targeted dianthin is a powerful toxin to treat pancreatic carcinoma when applied in combination with the glycosylated triterpene SO1861.Mol Oncol. 2017 Nov;11(11):1527-1543. doi: 10.1002/1878-0261.12115. Epub 2017 Sep 15. Mol Oncol. 2017. PMID: 28755527 Free PMC article.
-
Improved Therapy of B-Cell Non-Hodgkin Lymphoma by Obinutuzumab-Dianthin Conjugates in Combination with the Endosomal Escape Enhancer SO1861.Toxins (Basel). 2022 Jul 13;14(7):478. doi: 10.3390/toxins14070478. Toxins (Basel). 2022. PMID: 35878216 Free PMC article.
-
[Highly toxic type Ⅱ ribosome-inactivating proteins ricin and abrin and their detection methods: a review].Se Pu. 2021 Mar;39(3):260-270. doi: 10.3724/SP.J.1123.2020.10001. Se Pu. 2021. PMID: 34227307 Free PMC article. Review. Chinese.
Cited by
-
Potentiation of Gelonin Cytotoxicity by Pulsed Electric Fields.Int J Mol Sci. 2025 Jan 8;26(2):458. doi: 10.3390/ijms26020458. Int J Mol Sci. 2025. PMID: 39859180 Free PMC article.
References
-
- Endo Y., Mitsui K., Motizuki M., Tsurugi K. The mechanism of action of ricin and related toxic lectins on eukaryotic ribosomes. The site and the characteristics of the modification in 28 S ribosomal RNA caused by the toxins. J. Biol. Chem. 1987;262:5908–5912. doi: 10.1016/S0021-9258(18)45660-8. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
