

Human pluripotent stem cell-derived kidney organoids reveal tubular epithelial pathobiology of heterozygous HNF1B-associated dysplastic kidney malformations
- PMID: 38788724
- PMCID: PMC11297557
- DOI: 10.1016/j.stemcr.2024.04.011
Human pluripotent stem cell-derived kidney organoids reveal tubular epithelial pathobiology of heterozygous HNF1B-associated dysplastic kidney malformations
Abstract
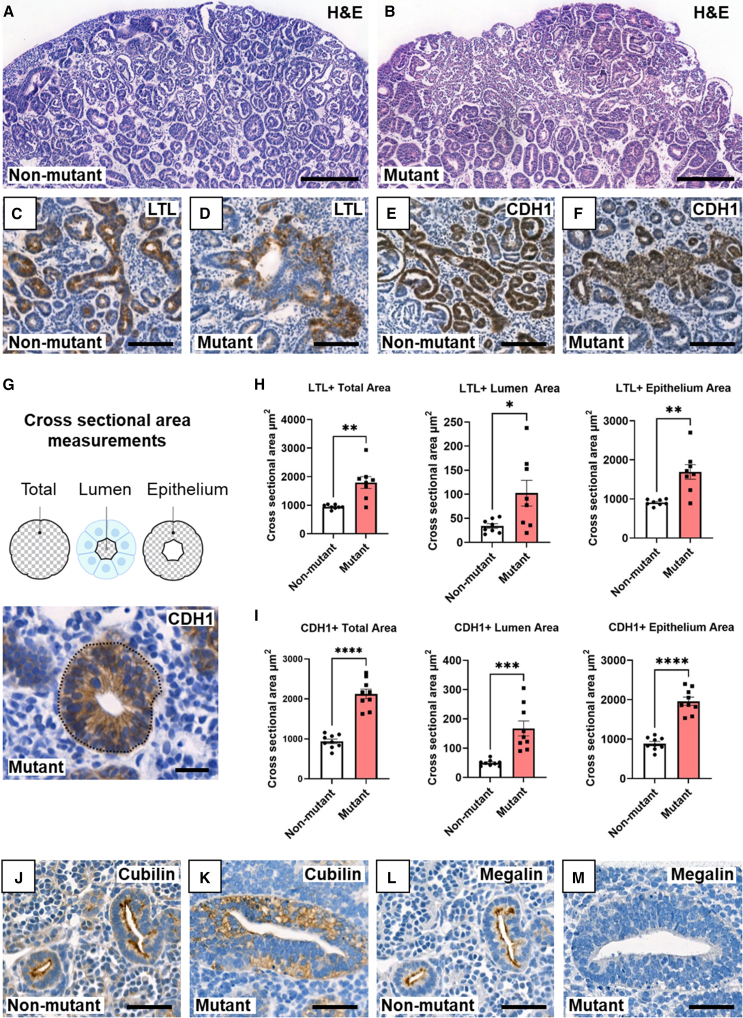
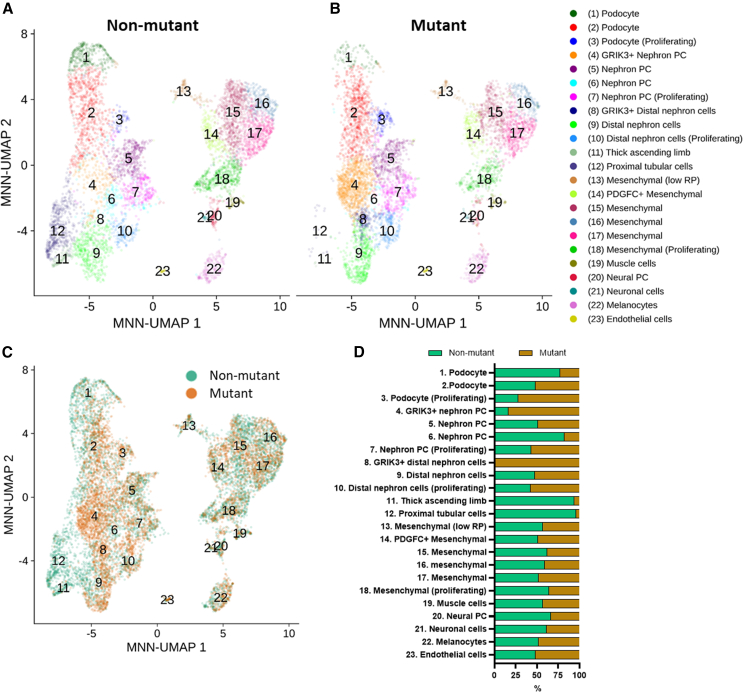
Hepatocyte nuclear factor 1B (HNF1B) encodes a transcription factor expressed in developing human kidney epithelia. Heterozygous HNF1B mutations are the commonest monogenic cause of dysplastic kidney malformations (DKMs). To understand their pathobiology, we generated heterozygous HNF1B mutant kidney organoids from CRISPR-Cas9 gene-edited human embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) reprogrammed from a family with HNF1B-associated DKMs. Mutant organoids contained enlarged malformed tubules displaying deregulated cell turnover. Numerous genes implicated in Mendelian kidney tubulopathies were downregulated, and mutant tubules resisted the cyclic AMP (cAMP)-mediated dilatation seen in controls. Bulk and single-cell RNA sequencing (scRNA-seq) analyses indicated abnormal Wingless/Integrated (WNT), calcium, and glutamatergic pathways, the latter hitherto unstudied in developing kidneys. Glutamate ionotropic receptor kainate type subunit 3 (GRIK3) was upregulated in malformed mutant nephron tubules and prominent in HNF1B mutant fetal human dysplastic kidney epithelia. These results reveal morphological, molecular, and physiological roles for HNF1B in human kidney tubule differentiation and morphogenesis illuminating the developmental origin of mutant-HNF1B-causing kidney disease.
Keywords: CRISPR; GRIK3; HNF1B; cAMP; glutamate receptors; kidney RNA-seq; kidney disease; organoid; pluripotent stem cells; proximal tubule.
Crown Copyright © 2024. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflicts of interest.
Figures






References
-
- Adalat S., Hayes W.N., Bryant W.A., Booth J., Woolf A.S., Kleta R., Subtil S., Clissold R., Colclough K., Ellard S., Bockenhauer D. HNF1B Mutations Are Associated With a Gitelman-like Tubulopathy That Develops During Childhood. Kidney Int. Rep. 2019;4:1304–1311. doi: 10.1016/j.ekir.2019.05.019. - DOI - PMC - PubMed
-
- Adalat S., Woolf A.S., Johnstone K.A., Wirsing A., Harries L.W., Long D.A., Hennekam R.C., Ledermann S.E., Rees L., van't Hoff W., et al. HNF1B mutations associate with hypomagnesemia and renal magnesium wasting. J. Am. Soc. Nephrol. 2009;20:1123–1131. doi: 10.1681/ASN.2008060633. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
