

The Activity of YCA1 Metacaspase Is Regulated by Reactive Sulfane Sulfur via Persulfidation in Saccharomyces cerevisiae
- PMID: 38790694
- PMCID: PMC11118234
- DOI: 10.3390/antiox13050589
The Activity of YCA1 Metacaspase Is Regulated by Reactive Sulfane Sulfur via Persulfidation in Saccharomyces cerevisiae
Abstract
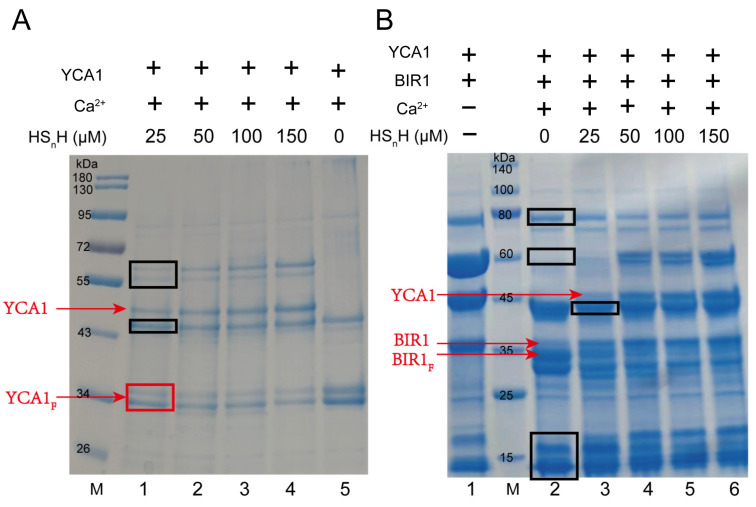
YCA1, the only metacaspase in Saccharomyces cerevisiae, plays important roles in the regulation of chronological lifespan, apoptosis, and cytokinesis. YCA1 has protein hydrolase activity and functions by cleaving itself and target proteins. However, there are few reports about the regulation of YCA1 activity. In this study, we observed that reactive sulfane sulfur (RSS) can inhibit the activity of YCA1. In vitro experiments demonstrated that RSS reacted with the Cys276 of YCA1, the residue central to its protein hydrolase activity, to form a persulfidation modification (protein-SSH). This modification inhibited both its self-cleavage and the cleavage of its substrate protein, BIR1. To investigate further, we constructed a low-endogenous-RSS mutant of S. cerevisiae, BY4742 Δcys3, in which the RSS-producing enzyme cystathionine-γ-lyase (CYS3) was knocked out. The activity of YCA1 was significantly increased by the deletion of CYS3. Moreover, increased YCA1 activity led to reduced chronological lifespan (CLS) and CLS-driven apoptosis. This study unveils the first endogenous factor that regulates YCA1 activity, introduces a novel mechanism of how yeast cells regulate chronological lifespan, and broadens our understanding of the multifaceted roles played by RSS.
Keywords: YCA1; apoptosis; chronological lifespan; metacaspase; persulfidation; reactive sulfane sulfur.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures





References
Grants and funding
LinkOut - more resources
Full Text Sources
