

Antibiotic Loaded Phytosomes as a Way to Develop Innovative Lipid Formulations of Polyene Macrolides
- PMID: 38794328
- PMCID: PMC11124810
- DOI: 10.3390/pharmaceutics16050665
Antibiotic Loaded Phytosomes as a Way to Develop Innovative Lipid Formulations of Polyene Macrolides
Abstract
Background: The threat of antibiotic resistance of fungal pathogens and the high toxicity of the most effective drugs, polyene macrolides, force us to look for new ways to develop innovative antifungal formulations.
Objective: The aim of this study was to determine how the sterol, phospholipid, and flavonoid composition of liposomal forms of polyene antibiotics, and in particular, amphotericin B (AmB), affects their ability to increase the permeability of lipid bilayers that mimic the membranes of mammalian and fungal cells.
Methods: To monitor the membrane permeability induced by various polyene-based lipid formulations, a calcein leakage assay and the electrophysiological technique based on planar lipid bilayers were used.
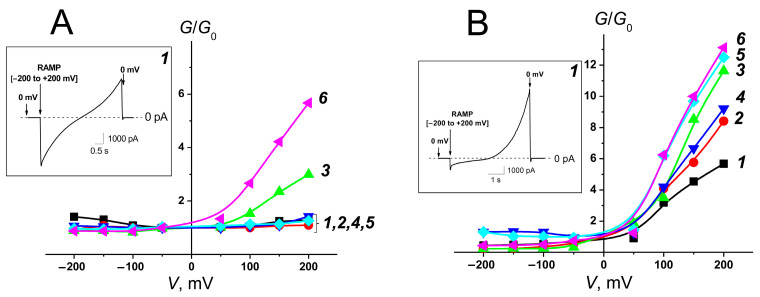
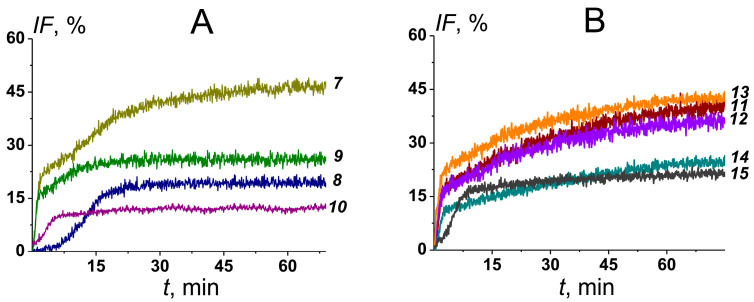
Key results: The replacement of cholesterol with its biosynthetic precursor, 7-dehydrocholesterol, led to a decrease in the ability of AmB-loaded liposomes to permeabilize lipid bilayers mimicking mammalian cell membranes. The inclusion of plant flavonoid phloretin in AmB-loaded liposomes increased the ability of the formulation to disengage a fluorescent marker from lipid vesicles mimicking the membranes of target fungi. I-V characteristics of the fungal-like lipid bilayers treated with the AmB phytosomes were symmetric, demonstrating the functioning of double-length AmB pores and assuming a decrease in the antibiotic threshold concentration.
Conclusions and perspectives: The therapeutic window of polyene lipid formulations might be expanded by varying their sterol composition. Polyene-loaded phytosomes might be considered as the prototypes for innovative lipid antibiotic formulations.
Keywords: amphotericin B; lipid bilayers; liposomes; membranes; phytosomes; sterols.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures





Similar articles
-
The structuring effects of amphotericin B on pure and ergosterol- or cholesterol-containing dipalmitoylphosphatidylcholine bilayers: a differential scanning calorimetry study.Biochim Biophys Acta. 1998 Aug 14;1373(1):76-86. doi: 10.1016/s0005-2736(98)00083-2. Biochim Biophys Acta. 1998. PMID: 9733926
-
How do the polyene macrolide antibiotics affect the cellular membrane properties?Biochim Biophys Acta. 1986 Dec 22;864(3-4):257-304. doi: 10.1016/0304-4157(86)90002-x. Biochim Biophys Acta. 1986. PMID: 3539192 Review.
-
Triggering the Amphotericin B Pore-Forming Activity by Phytochemicals.Membranes (Basel). 2023 Jul 14;13(7):670. doi: 10.3390/membranes13070670. Membranes (Basel). 2023. PMID: 37505036 Free PMC article.
-
How do sterols determine the antifungal activity of amphotericin B? Free energy of binding between the drug and its membrane targets.J Am Chem Soc. 2010 Dec 29;132(51):18266-72. doi: 10.1021/ja1074344. Epub 2010 Dec 2. J Am Chem Soc. 2010. PMID: 21126070
-
Liposomal amphotericin B: a review of its use as empirical therapy in febrile neutropenia and in the treatment of invasive fungal infections.Drugs. 2009;69(3):361-92. doi: 10.2165/00003495-200969030-00010. Drugs. 2009. PMID: 19275278 Review.
References
-
- de Kruijff B., Gerritsen W.J., Oerlemans A., Demel R.A., van Deenen L.L. Polyene antibiotic-sterol interactions in membranes of Acholeplasma laidlawii cells and lecithin liposomes. I. Specificity of the membrane permeability changes induced by the polyene antibiotics. Biochim. Biophys. Acta. 1974;339:30–43. doi: 10.1016/0005-2736(74)90330-7. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
