

Gut bacteria convert glucocorticoids into progestins in the presence of hydrogen gas
- PMID: 38795705
- PMCID: PMC11179439
- DOI: 10.1016/j.cell.2024.05.005
Gut bacteria convert glucocorticoids into progestins in the presence of hydrogen gas
Abstract
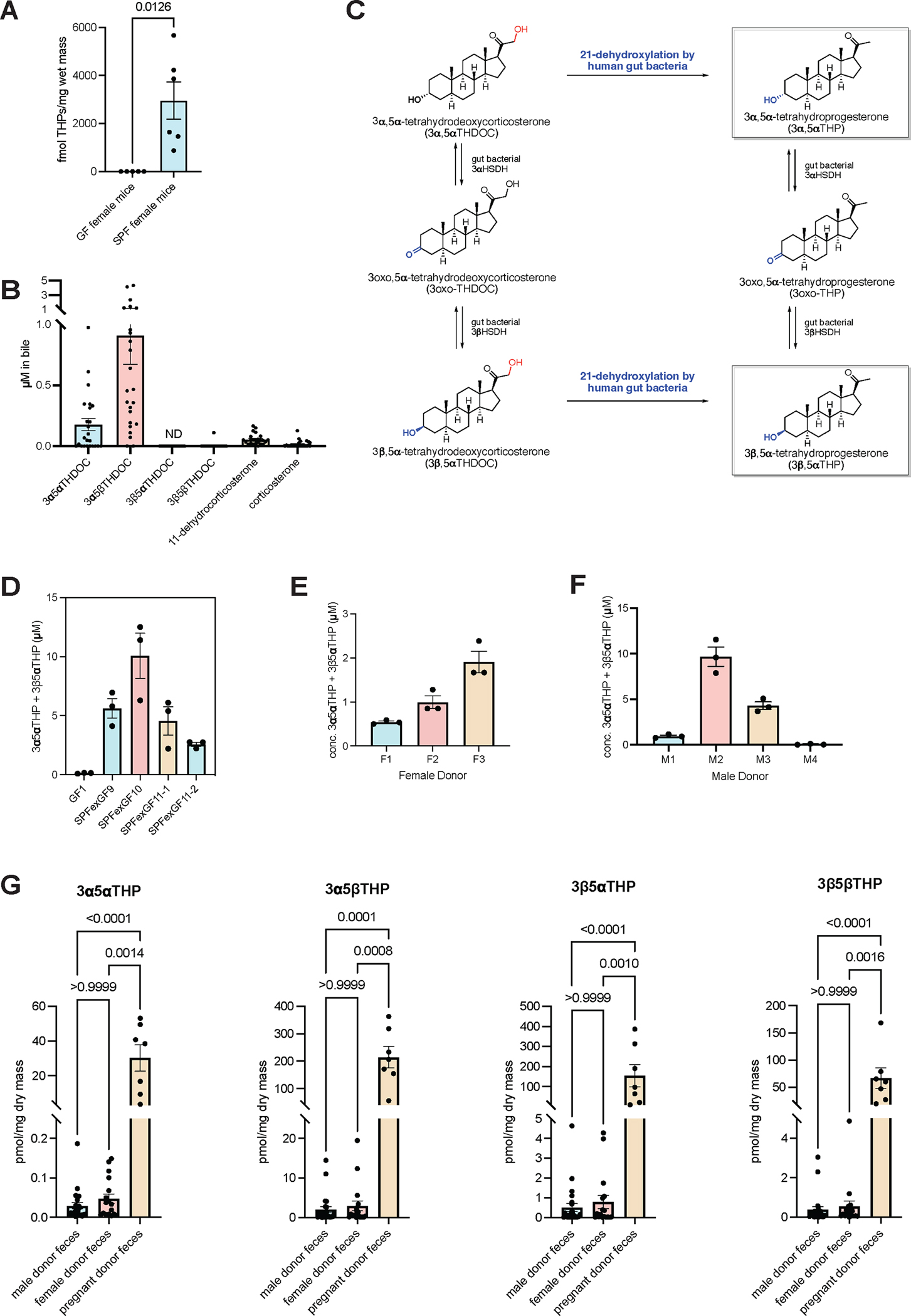
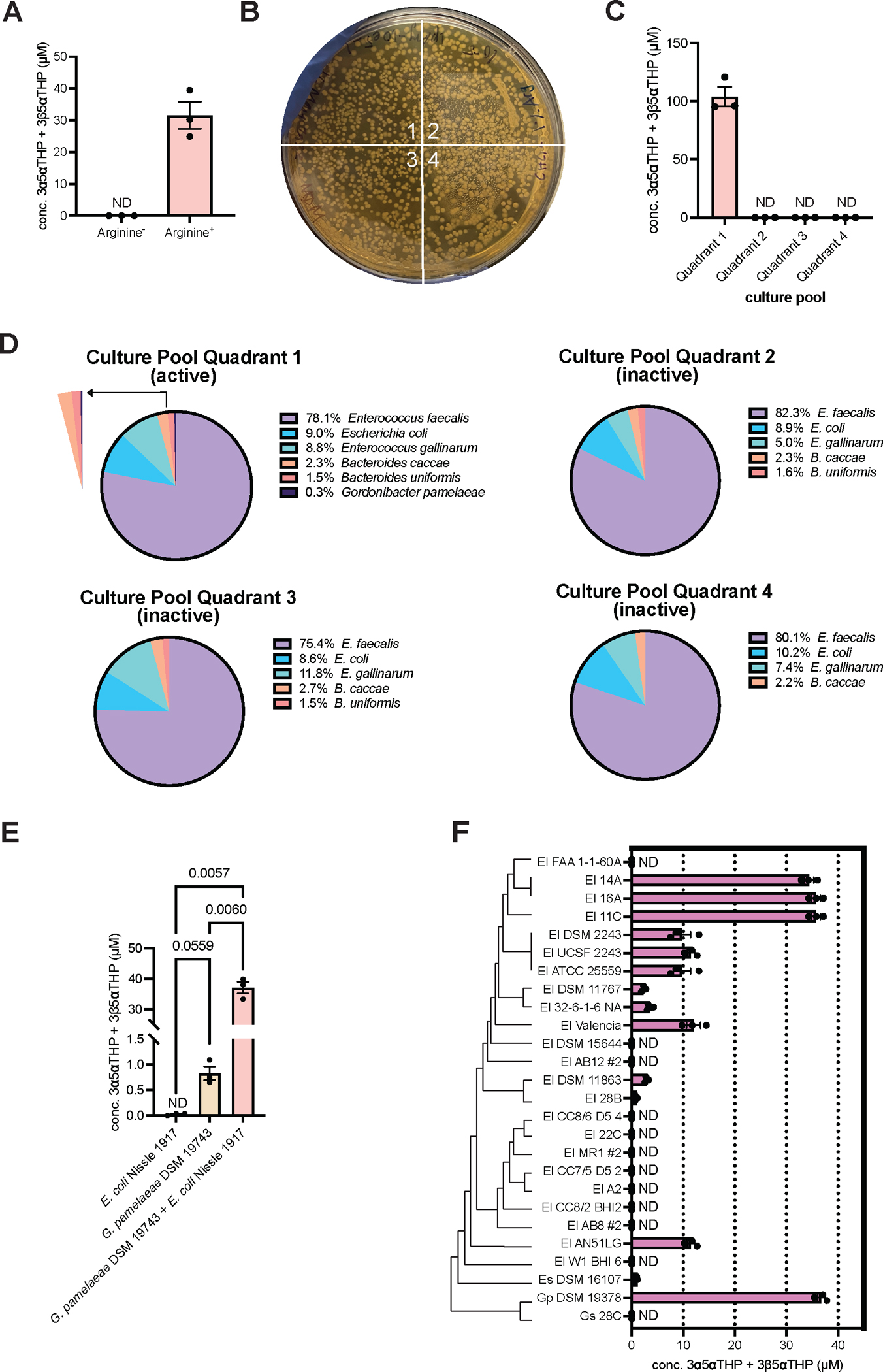
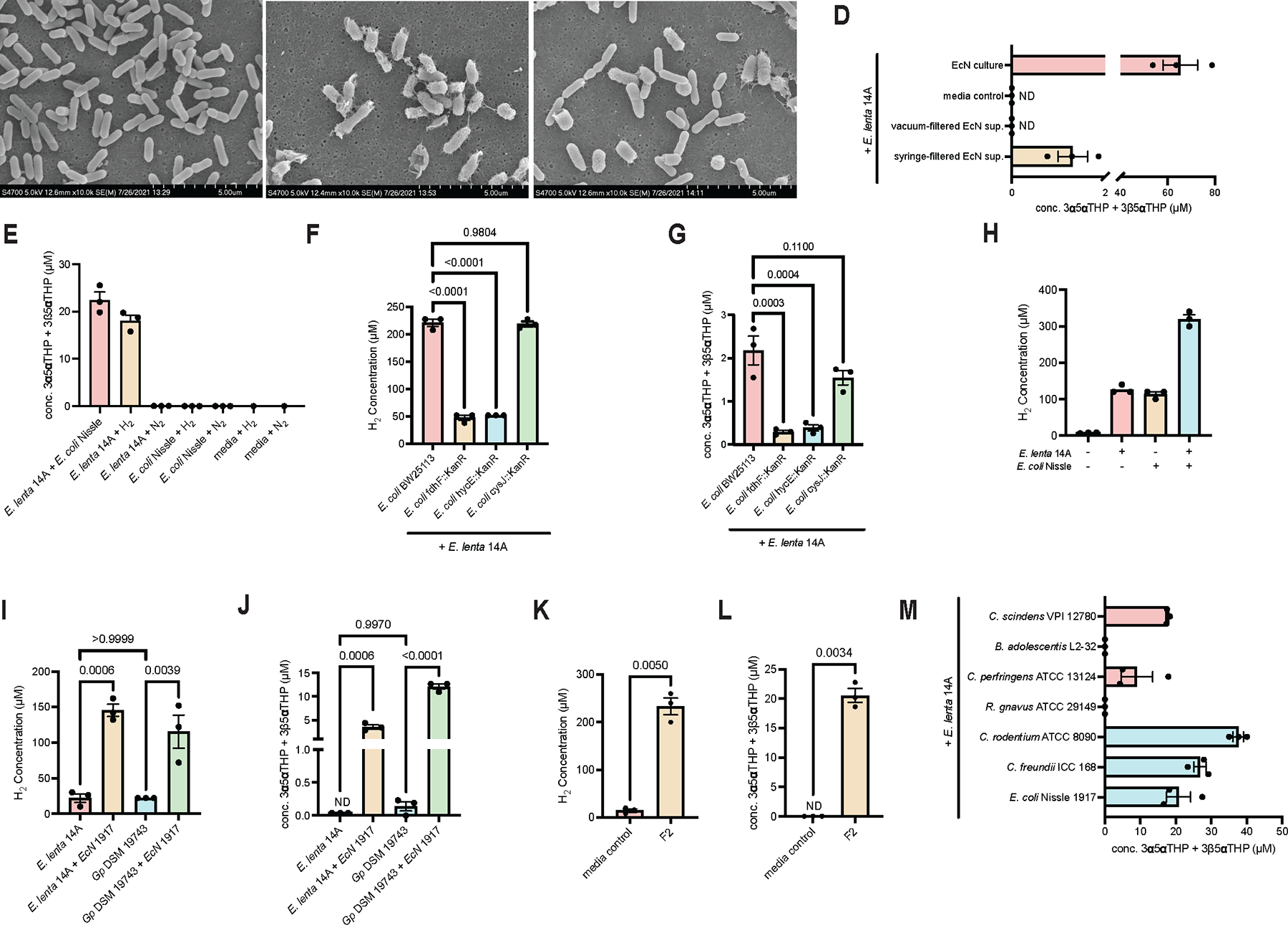
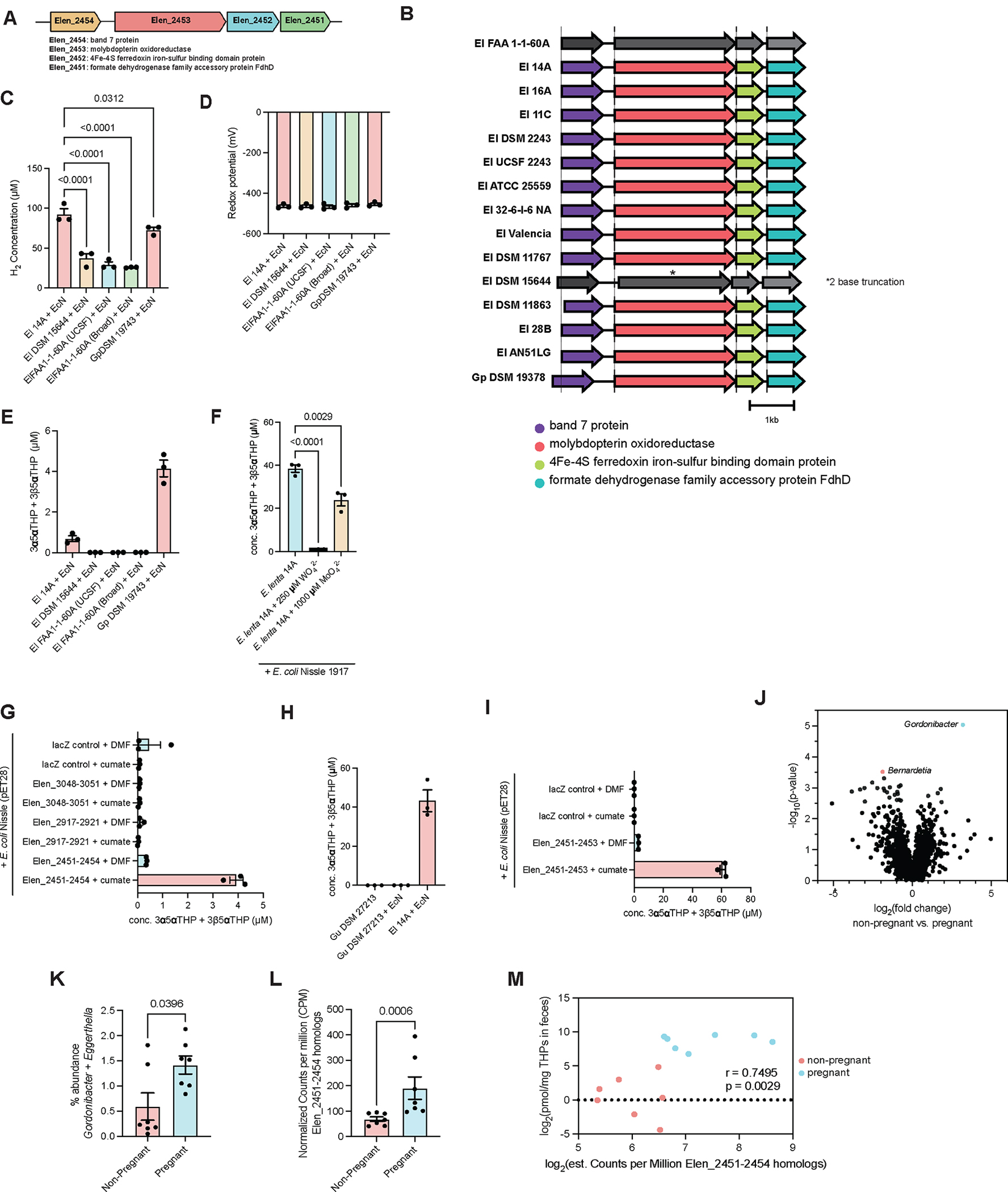
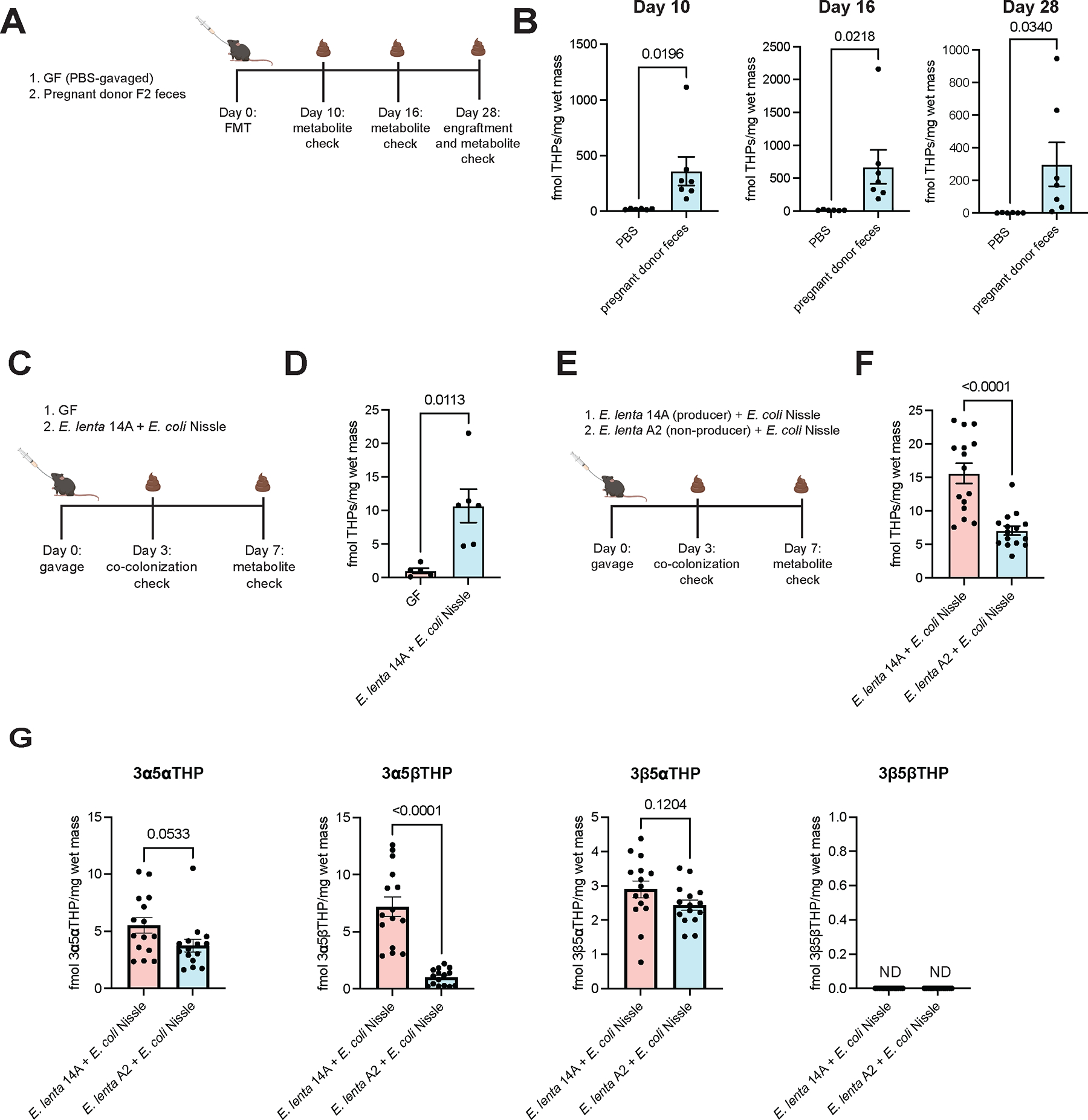
Recent studies suggest that human-associated bacteria interact with host-produced steroids, but the mechanisms and physiological impact of such interactions remain unclear. Here, we show that the human gut bacteria Gordonibacter pamelaeae and Eggerthella lenta convert abundant biliary corticoids into progestins through 21-dehydroxylation, thereby transforming a class of immuno- and metabo-regulatory steroids into a class of sex hormones and neurosteroids. Using comparative genomics, homologous expression, and heterologous expression, we identify a bacterial gene cluster that performs 21-dehydroxylation. We also uncover an unexpected role for hydrogen gas production by gut commensals in promoting 21-dehydroxylation, suggesting that hydrogen modulates secondary metabolism in the gut. Levels of certain bacterial progestins, including allopregnanolone, better known as brexanolone, an FDA-approved drug for postpartum depression, are substantially increased in feces from pregnant humans. Thus, bacterial conversion of corticoids into progestins may affect host physiology, particularly in the context of pregnancy and women's health.
Keywords: 21-dehydroxylation; Eggerthella lenta; hydrogen gas; neurosteroids.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.S.D. is an ad hoc consultant for Axial Therapeutics. M.D.M. and A.S.D. are co-inventors on a provisional patent related to this work. J.R.H. serves as a consultant for CJ CheilJedang and is a co-founder and consultant for Interon Laboratories. He received research funding from CJ Bioscience outside the submitted work. A.G.E. serves as a consultant for Mirvie, Inc. outside the submitted work and receives research funding from Merck Pharmaceuticals outside the submitted work.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
