

This is a preprint.
Single-cell morphology encodes functional subtypes of senescence in aging human dermal fibroblasts
- PMID: 38798365
- PMCID: PMC11118441
- DOI: 10.1101/2024.05.10.593637
Single-cell morphology encodes functional subtypes of senescence in aging human dermal fibroblasts
Update in
-
Single-cell morphology encodes functional subtypes of senescence in aging human dermal fibroblasts.Sci Adv. 2025 Apr 25;11(17):eads1875. doi: 10.1126/sciadv.ads1875. Epub 2025 Apr 25. Sci Adv. 2025. PMID: 40279419 Free PMC article.
Abstract
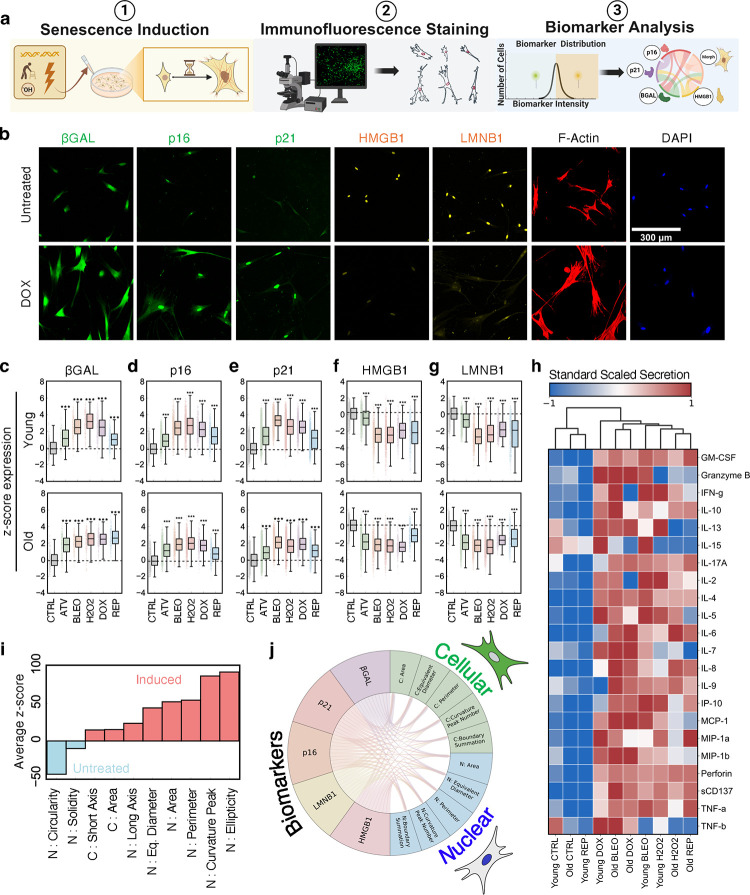
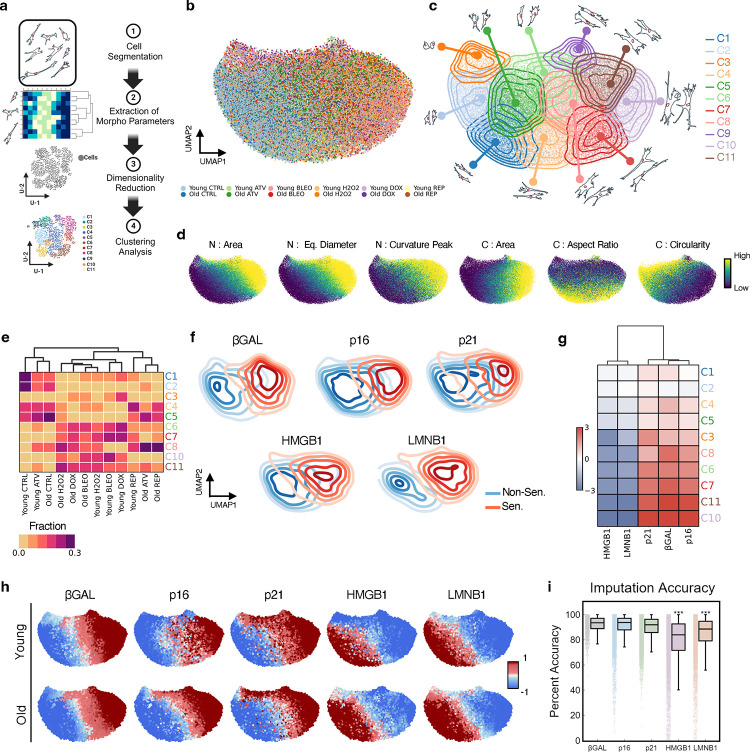
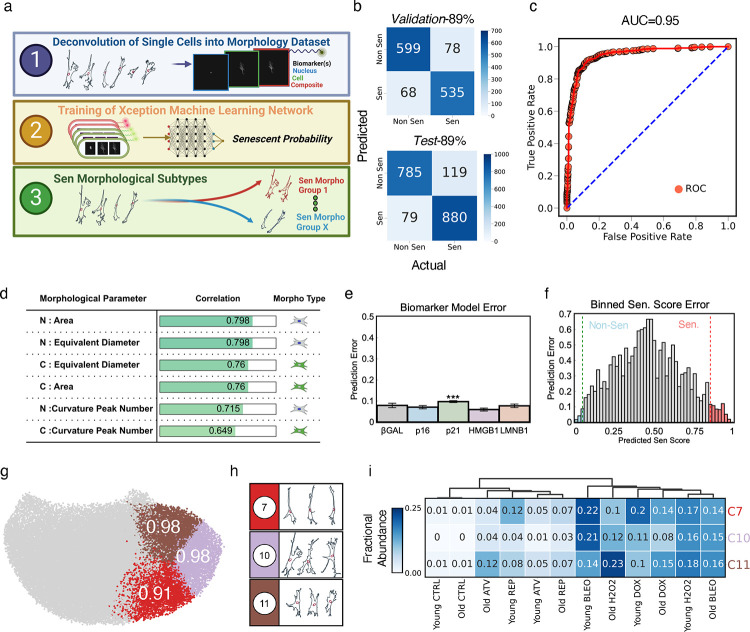
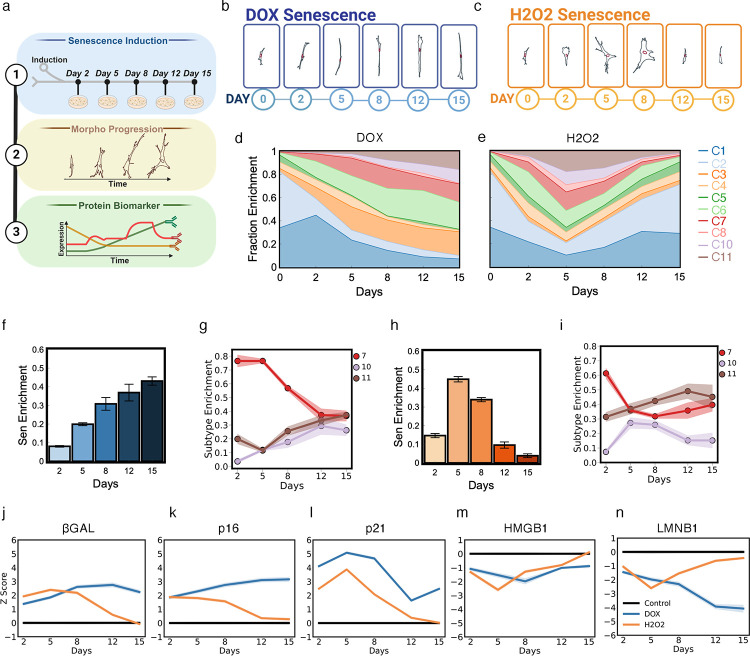
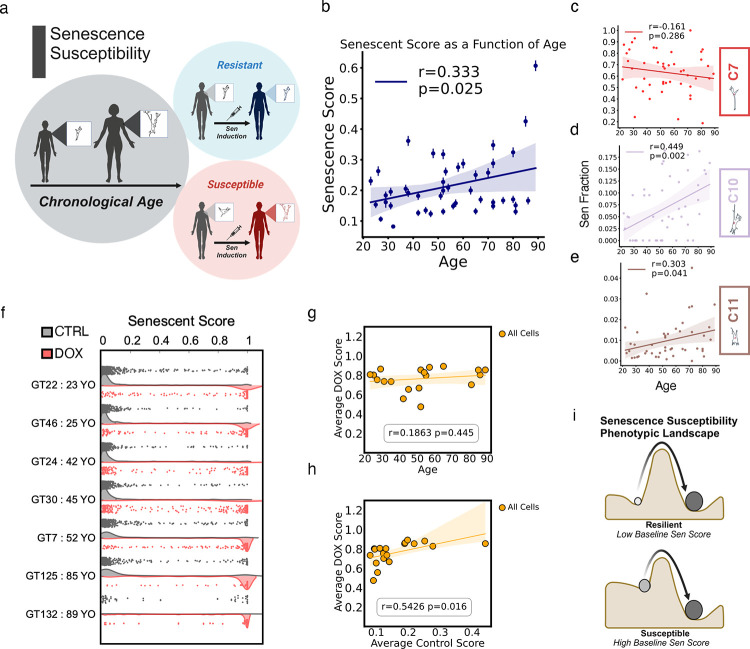
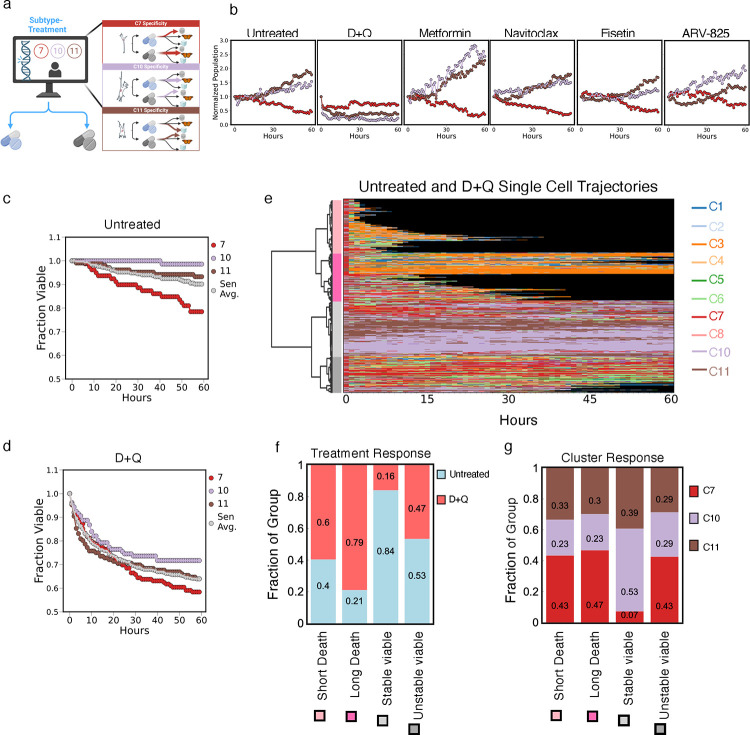
Cellular senescence is an established driver of aging, exhibiting context-dependent phenotypes across multiple biological length-scales. Despite its mechanistic importance, profiling senescence within cell populations is challenging. This is in part due to the limitations of current biomarkers to robustly identify senescent cells across biological settings, and the heterogeneous, non-binary phenotypes exhibited by senescent cells. Using a panel of primary dermal fibroblasts, we combined live single-cell imaging, machine learning, multiple senescence induction conditions, and multiple protein-based senescence biomarkers to show the emergence of functional subtypes of senescence. Leveraging single-cell morphologies, we defined eleven distinct morphology clusters, with the abundance of cells in each cluster being dependent on the mode of senescence induction, the time post-induction, and the age of the donor. Of these eleven clusters, we identified three bona-fide senescence subtypes (C7, C10, C11), with C10 showing the strongest age-dependence across a cohort of fifty aging individuals. To determine the functional significance of these senescence subtypes, we profiled their responses to senotherapies, specifically focusing on Dasatinib + Quercetin (D+Q). Results indicated subtype-dependent responses, with senescent cells in C7 being most responsive to D+Q. Altogether, we provide a robust single-cell framework to identify and classify functional senescence subtypes with applications for next-generation senotherapy screens, and the potential to explain heterogeneous senescence phenotypes across biological settings based on the presence and abundance of distinct senescence subtypes.
Keywords: aging; biomarkers; cell morphology; cellular senescence; functional subtypes.
Conflict of interest statement
Conflict of interest. P.K, N.M., J.M.P are inventors on a patent application related to this work. All other authors declare no conflict of interest.
Figures
References
-
- Hernandez-Segura A., Nehme J. & Demaria M. Hallmarks of Cellular Senescence. Trends Cell Biol 28, 436–453 (2018). - PubMed
-
- Nathan Basisty AND Kale A. A. N. D. J. O. H. A. N. D. K. C. A. N. D. P. T. A. N. D. R. C. A. N. D. H. A. A. N. D. S. S. A. N. D. S. V. A. N. D. F. L. A. N. D. C. J. A. N. D. S. B. A proteomic atlas of senescence-associated secretomes for aging biomarker development. PLoS Biol 18, 1–26 (2020). - PMC - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous