

A fluorescence-based sensor for calibrated measurement of protein kinase stability in live cells
- PMID: 38801214
- PMCID: PMC11129626
- DOI: 10.1002/pro.5023
A fluorescence-based sensor for calibrated measurement of protein kinase stability in live cells
Abstract
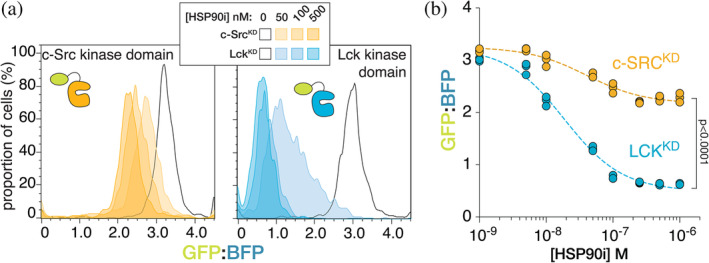
Oncogenic mutations can destabilize signaling proteins, resulting in increased or unregulated activity. Thus, there is considerable interest in mapping the relationship between mutations and the stability of signaling proteins, to better understand the consequences of oncogenic mutations and potentially inform the development of new therapeutics. Here, we develop a tool to study protein-kinase stability in live mammalian cells and the effects of the HSP90 chaperone system on the stability of these kinases. We determine the expression levels of protein kinases by monitoring the fluorescence of fluorescent proteins fused to those kinases, normalized to that of co-expressed reference fluorescent proteins. We used this tool to study the dependence of Src- and Raf-family kinases on the HSP90 system. We demonstrate that this sensor reports on destabilization induced by oncogenic mutations in these kinases. We also show that Src-homology 2 and Src-homology 3 domains, which are required for autoinhibition of Src-family kinases, stabilize these kinase domains in the cell. Our expression-calibrated sensor enables the facile characterization of the effects of mutations and small-molecule drugs on protein-kinase stability.
Keywords: HSP90; cell signaling; fluorescent sensors; kinase inhibitors; kinases; oncogenes; protein stability.
© 2024 The Authors. Protein Science published by Wiley Periodicals LLC on behalf of The Protein Society.
Conflict of interest statement
John Kuriyan is the Editor‐in‐Chief of
Figures




Update of
-
A Fluorescence-Based Sensor for Calibrated Measurement of Protein Kinase Stability in Live Cells.bioRxiv [Preprint]. 2023 Dec 8:2023.12.07.570636. doi: 10.1101/2023.12.07.570636. bioRxiv. 2023. Update in: Protein Sci. 2024 Jun;33(6):e5023. doi: 10.1002/pro.5023. PMID: 38106090 Free PMC article. Updated. Preprint.
Similar articles
-
A Fluorescence-Based Sensor for Calibrated Measurement of Protein Kinase Stability in Live Cells.bioRxiv [Preprint]. 2023 Dec 8:2023.12.07.570636. doi: 10.1101/2023.12.07.570636. bioRxiv. 2023. Update in: Protein Sci. 2024 Jun;33(6):e5023. doi: 10.1002/pro.5023. PMID: 38106090 Free PMC article. Updated. Preprint.
-
Hsp90 dependence of a kinase is determined by its conformational landscape.Sci Rep. 2017 Mar 14;7:43996. doi: 10.1038/srep43996. Sci Rep. 2017. PMID: 28290541 Free PMC article.
-
Hsp90 is essential for the synthesis and subsequent membrane association, but not the maintenance, of the Src-kinase p56(lck).Mol Biol Cell. 2000 May;11(5):1585-95. doi: 10.1091/mbc.11.5.1585. Mol Biol Cell. 2000. PMID: 10793137 Free PMC article.
-
Inhibition of cancer invasion and metastasis by targeting the molecular chaperone heat-shock protein 90.Anticancer Res. 2009 Mar;29(3):797-807. Anticancer Res. 2009. PMID: 19414312 Review.
-
The role of the hsp90-based chaperone system in signal transduction by nuclear receptors and receptors signaling via MAP kinase.Annu Rev Pharmacol Toxicol. 1997;37:297-326. doi: 10.1146/annurev.pharmtox.37.1.297. Annu Rev Pharmacol Toxicol. 1997. PMID: 9131255 Review.
Cited by
-
Action and therapeutic targets of folliculin interacting protein 1: a novel signaling mechanism in redox regulation.Front Cell Dev Biol. 2025 Mar 12;13:1523489. doi: 10.3389/fcell.2025.1523489. eCollection 2025. Front Cell Dev Biol. 2025. PMID: 40143966 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
