

Regulation of cell distancing in peri-plaque glial nets by Plexin-B1 affects glial activation and amyloid compaction in Alzheimer's disease
- PMID: 38802590
- PMCID: PMC11346591
- DOI: 10.1038/s41593-024-01664-w
Regulation of cell distancing in peri-plaque glial nets by Plexin-B1 affects glial activation and amyloid compaction in Alzheimer's disease
Abstract
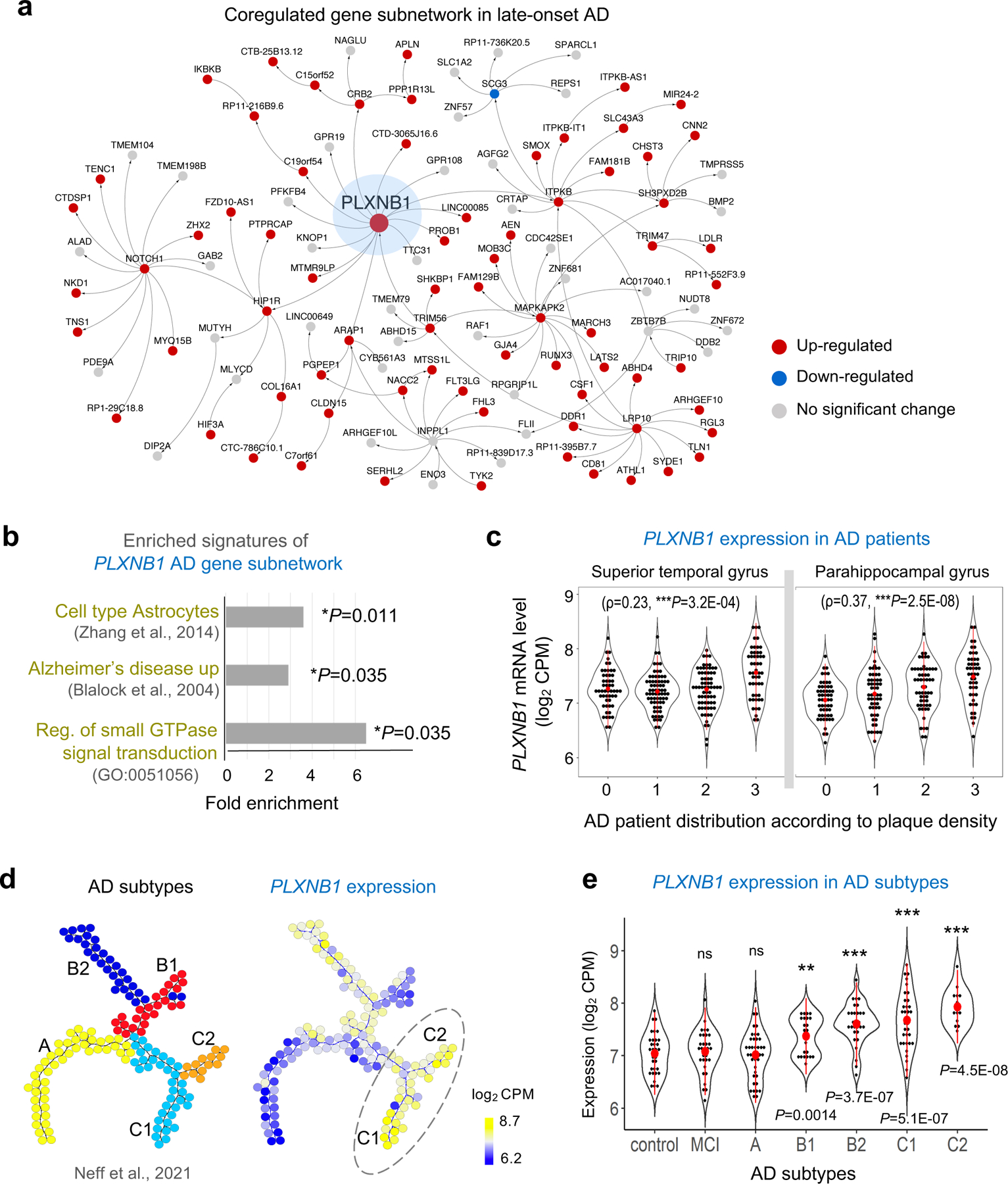
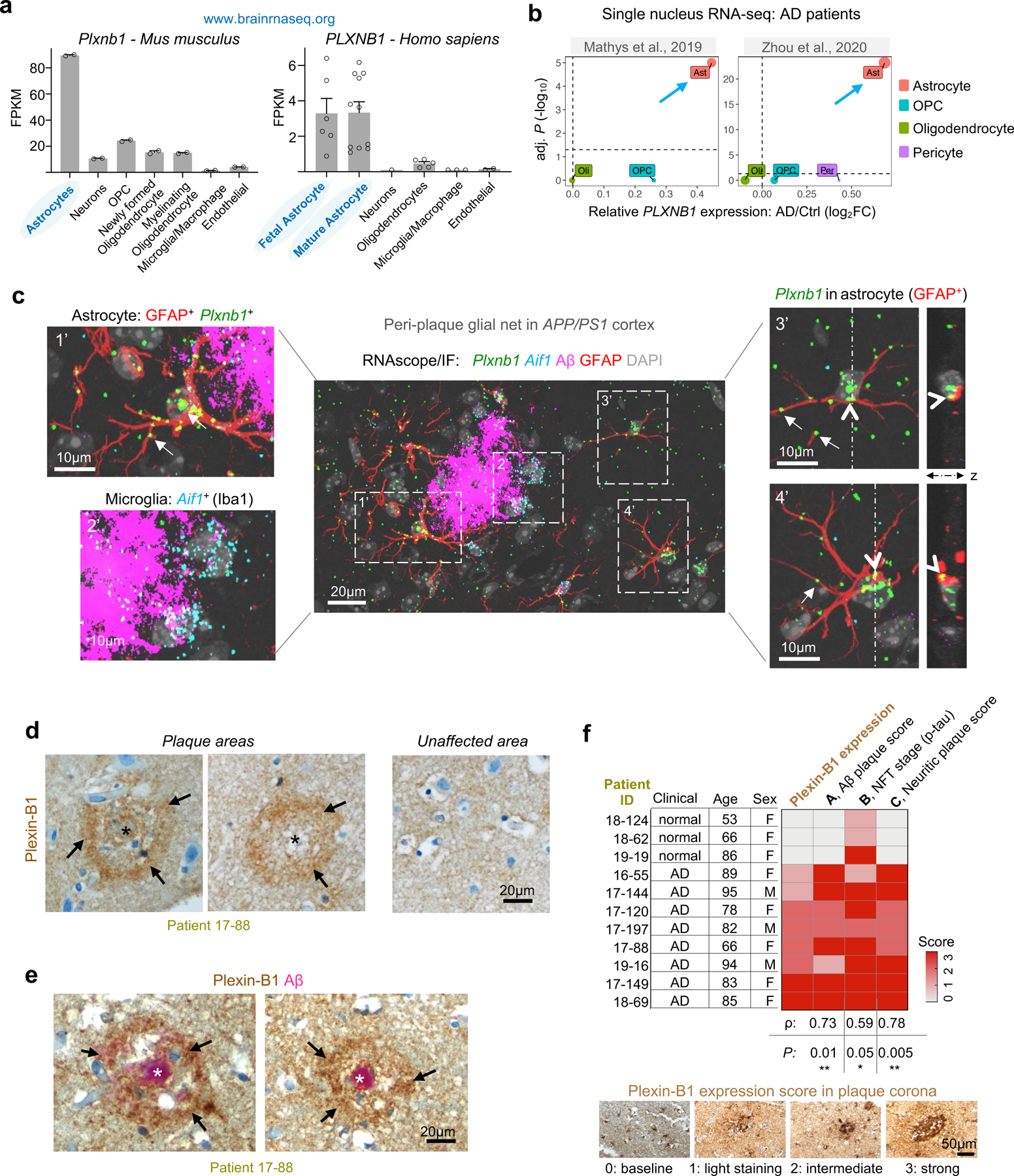
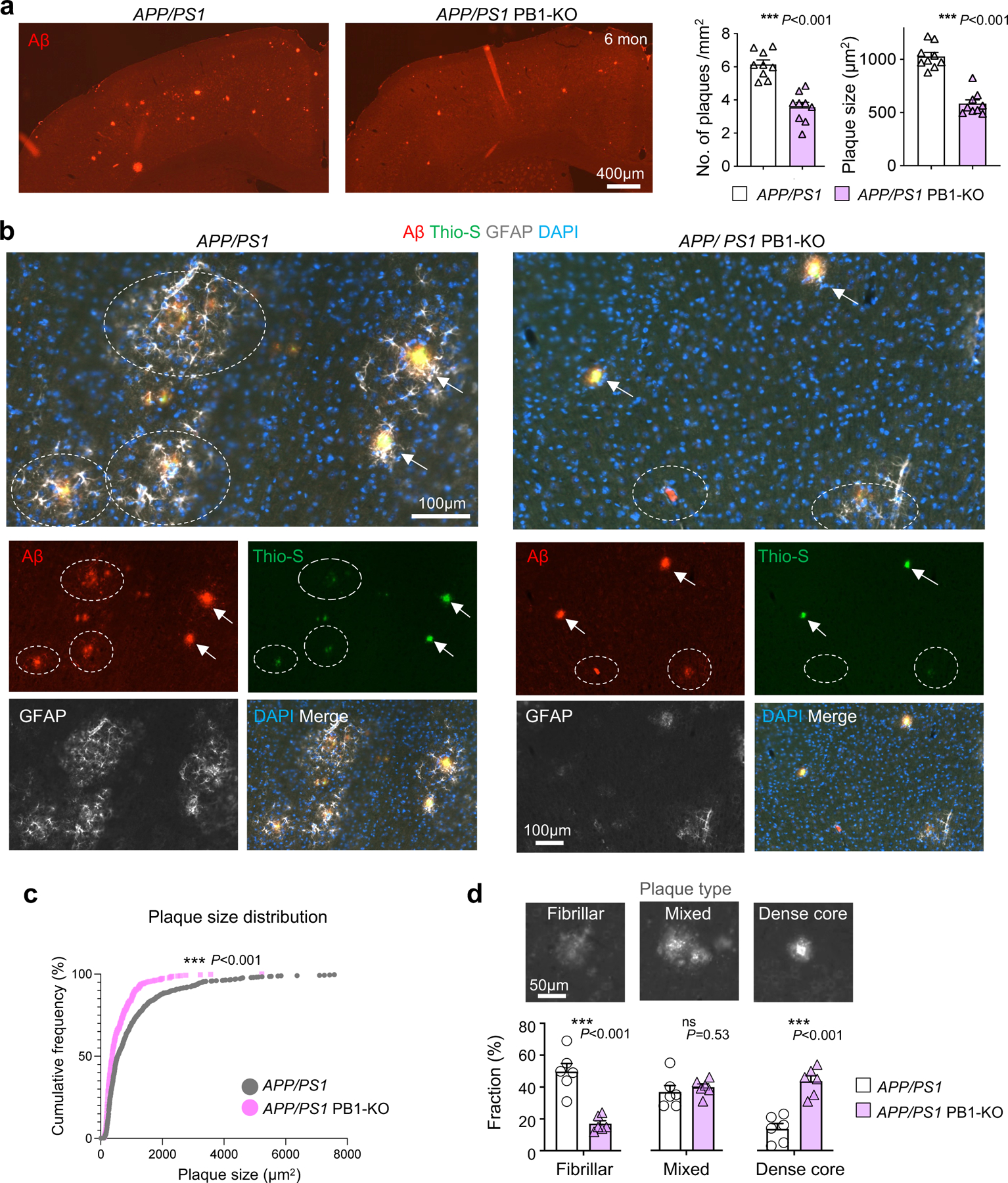
Communication between glial cells has a profound impact on the pathophysiology of Alzheimer's disease (AD). We reveal here that reactive astrocytes control cell distancing in peri-plaque glial nets, which restricts microglial access to amyloid deposits. This process is governed by guidance receptor Plexin-B1 (PLXNB1), a network hub gene in individuals with late-onset AD that is upregulated in plaque-associated astrocytes. Plexin-B1 deletion in a mouse AD model led to reduced number of reactive astrocytes and microglia in peri-plaque glial nets, but higher coverage of plaques by glial processes, along with transcriptional changes signifying reduced neuroinflammation. Additionally, a reduced footprint of glial nets was associated with overall lower plaque burden, a shift toward dense-core-type plaques and reduced neuritic dystrophy. Altogether, our study demonstrates that Plexin-B1 regulates peri-plaque glial net activation in AD. Relaxing glial spacing by targeting guidance receptors may present an alternative strategy to increase plaque compaction and reduce neuroinflammation in AD.
© 2024. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing Interests
The authors declare no competing interests.
Figures





References
-
- Leng F & Edison P Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here? Nat Rev Neurol 17, 157–172 (2021). - PubMed
Figure Legends References
MeSH terms
Substances
Grants and funding
- RF1 AG057440/AG/NIA NIH HHS/United States
- U01 AG046170/AG/NIA NIH HHS/United States
- R21 AG077168/AG/NIA NIH HHS/United States
- T32 CA078207/CA/NCI NIH HHS/United States
- RF1 AG077828/AG/NIA NIH HHS/United States
- RF1AG057440/U.S. Department of Health & Human Services | NIH | National Institute on Aging (U.S. National Institute on Aging)
- SCIRB C39068GG/New York State Department of Health (NYS Department of Health)
- U01AG046170/U.S. Department of Health & Human Services | NIH | National Institute on Aging (U.S. National Institute on Aging)
- R01 AG068030/AG/NIA NIH HHS/United States
- T32 GM146636/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
