

The circadian clock in the choroid plexus drives rhythms in multiple cellular processes under the control of the suprachiasmatic nucleus
- PMID: 38802875
- PMCID: PMC11131265
- DOI: 10.1186/s12987-024-00547-3
The circadian clock in the choroid plexus drives rhythms in multiple cellular processes under the control of the suprachiasmatic nucleus
Abstract
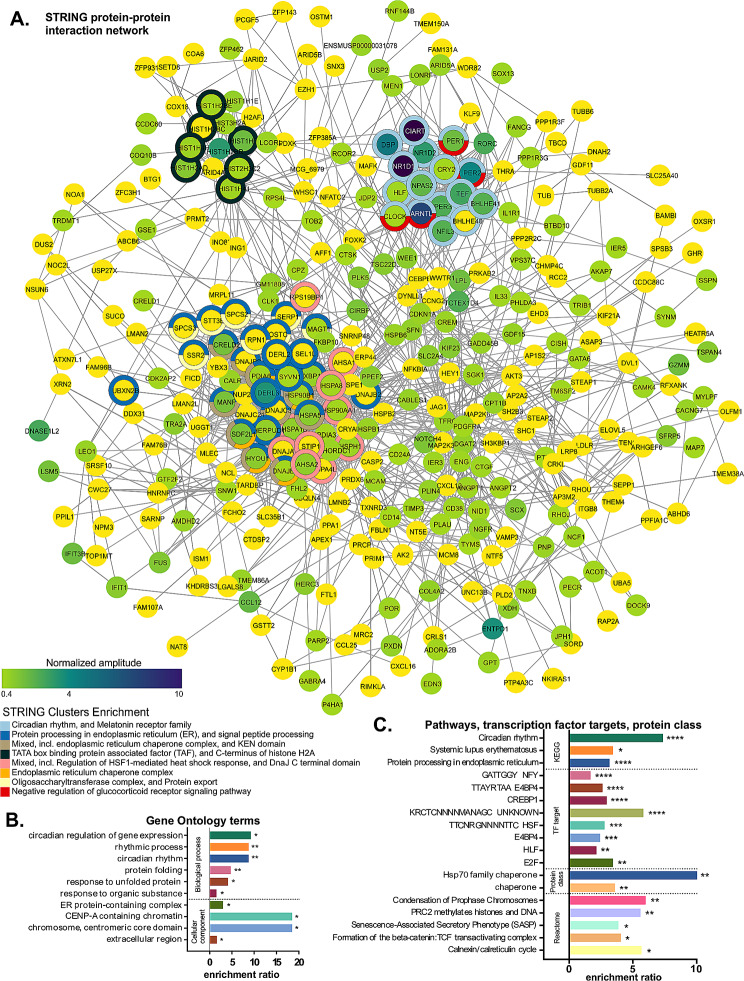
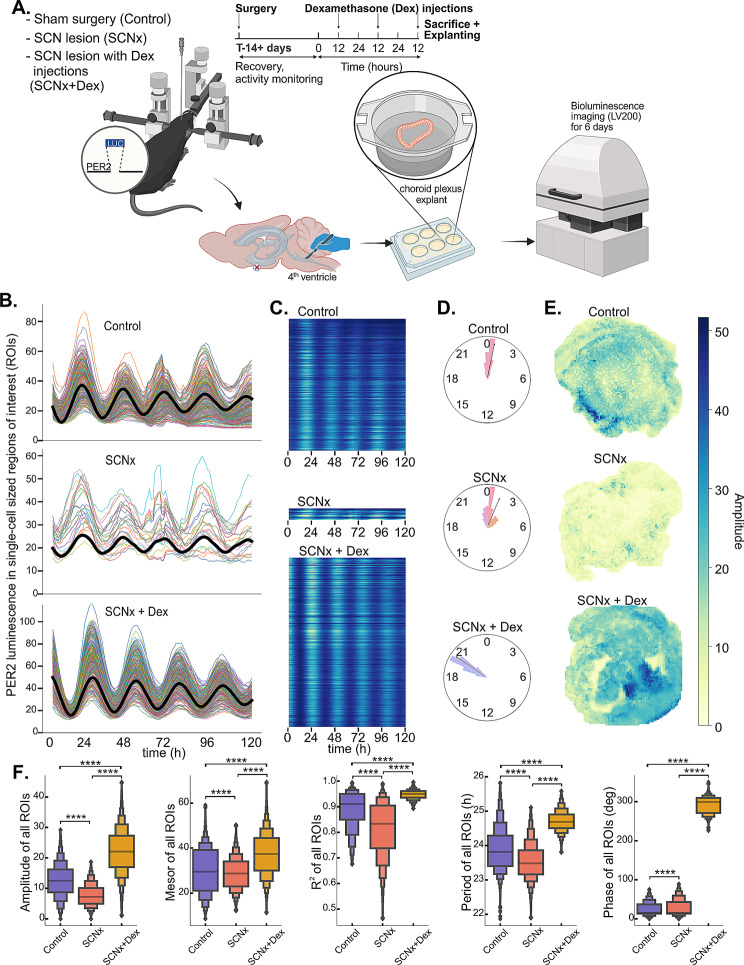
Choroid plexus (ChP), the brain structure primarily responsible for cerebrospinal fluid production, contains a robust circadian clock, whose role remains to be elucidated. The aim of our study was to [1] identify rhythmically controlled cellular processes in the mouse ChP and [2] assess the role and nature of signals derived from the master clock in the suprachiasmatic nuclei (SCN) that control ChP rhythms. To accomplish this goal, we used various mouse models (WT, mPer2Luc, ChP-specific Bmal1 knockout) and combined multiple experimental approaches, including surgical lesion of the SCN (SCNx), time-resolved transcriptomics, and single cell luminescence microscopy. In ChP of control (Ctrl) mice collected every 4 h over 2 circadian cycles in darkness, we found that the ChP clock regulates many processes, including the cerebrospinal fluid circadian secretome, precisely times endoplasmic reticulum stress response, and controls genes involved in neurodegenerative diseases (Alzheimer's disease, Huntington's disease, and frontotemporal dementia). In ChP of SCNx mice, the rhythmicity detected in vivo and ex vivo was severely dampened to a comparable extent as in mice with ChP-specific Bmal1 knockout, and the dampened cellular rhythms were restored by daily injections of dexamethasone in mice. Our data demonstrate that the ChP clock controls tissue-specific gene expression and is strongly dependent on the presence of a functional connection with the SCN. The results may contribute to the search for a novel link between ChP clock disruption and impaired brain health.
Keywords: mPer2 Luc mouse; Choroid plexus; Circadian clock; Circadian transcriptome; Glucocorticoid; Mouse; Suprachiasmatic nuclei.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Rennels ML, Gregory TF, Blaumanis OR, Fujimoto K, Grady PA. Evidence for a ‘paravascular’ fluid circulation in the mammalian central nervous system, provided by the rapid distribution of tracer protein throughout the brain from the subarachnoid space. Brain Res. 1985;326(1):47–63. doi: 10.1016/0006-8993(85)91383-6. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
