

Erwin Bünning and Wolfgang Engelmann: establishing the involvement of the circadian clock in photoperiodism
- PMID: 38805044
- PMCID: PMC11226508
- DOI: 10.1007/s00359-024-01704-7
Erwin Bünning and Wolfgang Engelmann: establishing the involvement of the circadian clock in photoperiodism
Abstract
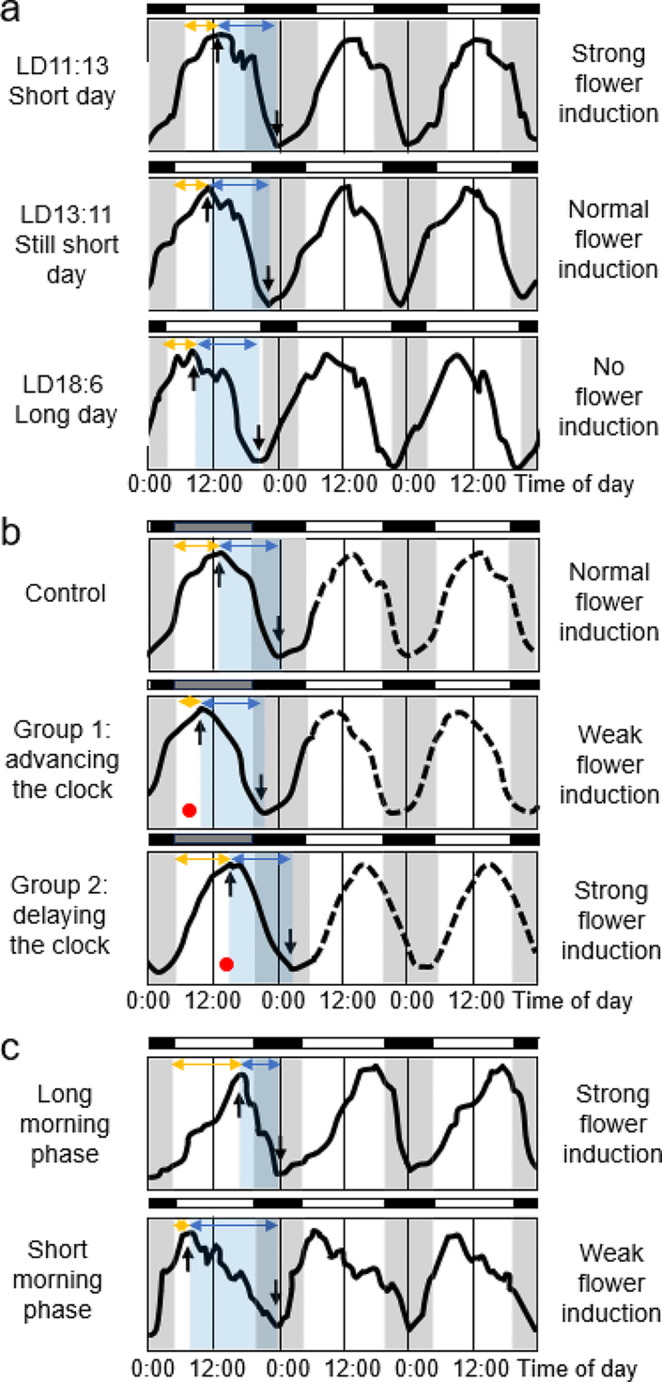
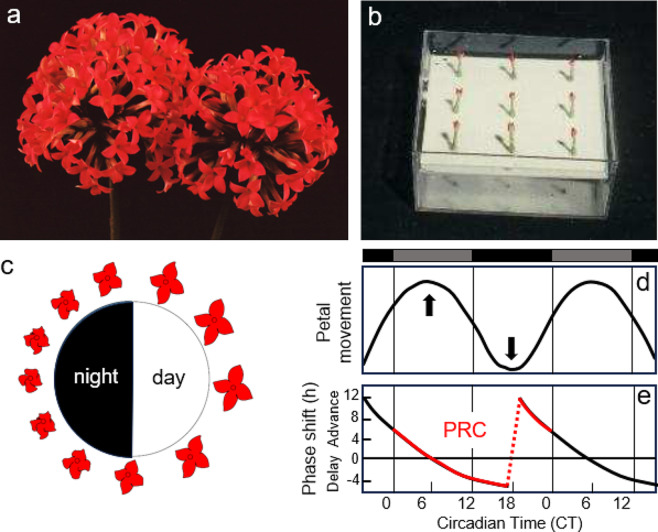
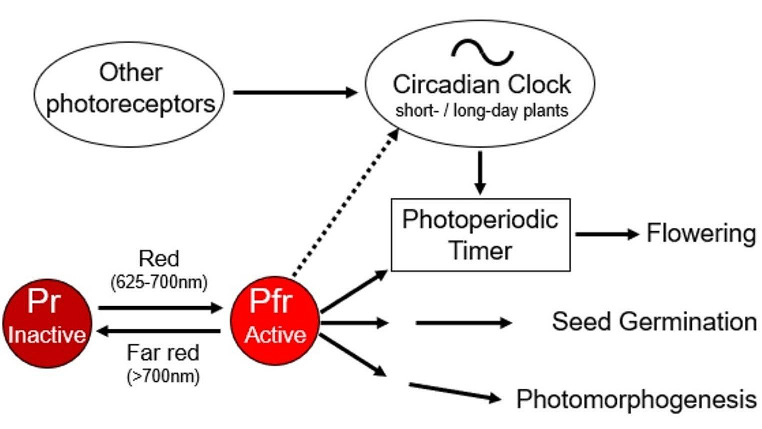
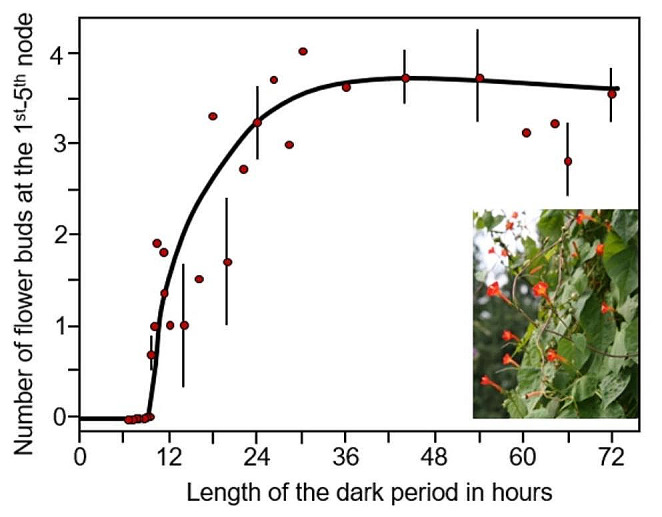
In 1936, Erwin Bünning published his groundbreaking work that the endogenous clock is used to measure day length for initiating photoperiodic responses. His publication triggered years of controversial debate until it ultimately became the basic axiom of rhythm research and the theoretical pillar of chronobiology. Bünning's thesis is frequently quoted in the articles in this special issue on the subject of "A clock for all seasons". However, nowadays only few people know in detail about Bünning's experiments and almost nobody knows about the contribution of his former doctoral student, Wolfgang Engelmann, to his theory because most work on this topic is published in German. The aim of this review is to give an overview of the most important experiments at that time, including Wolfgang Engelmann's doctoral thesis, in which he demonstrated the importance of the circadian clock for photoperiodic flower induction in the Flaming Katy, Kalanchoë blossfeldiana, but not in the Red Morning Glory, Ipomoea coccinea.
Keywords: Kalanchoë blossfeldiana; Phaseolus multiflorus; Circadian clock; Erwin Bünning; Photoperiodism; Wolfgang Engelmann.
© 2024. The Author(s).
Conflict of interest statement
CHF declares that she is editor of the Journal of Comparative Physiology A.
Figures


References
-
- Aksenova NP, Milyaeva EL, Romanov GA. Florigen goes molecular: seventy years of the hormonal theory of flowering regulation. Russ J Plant Physiol. 2006;33:401–406. doi: 10.1134/S1021443706030174. - DOI
-
- Beling I. Über das Zeitgedächtnis der Bienen. Z Vergl Physiol. 1929;9:259–338. doi: 10.1007/BF00340159. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
