

An essential role for EROS in redox-dependent endothelial signal transduction
- PMID: 38805973
- PMCID: PMC11153901
- DOI: 10.1016/j.redox.2024.103214
An essential role for EROS in redox-dependent endothelial signal transduction
Abstract
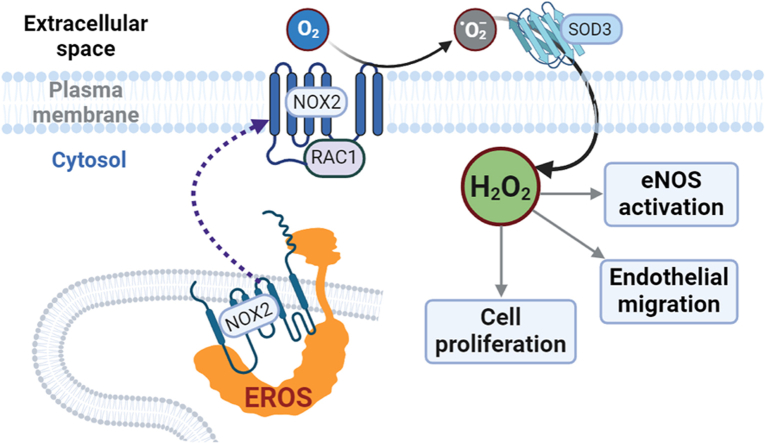
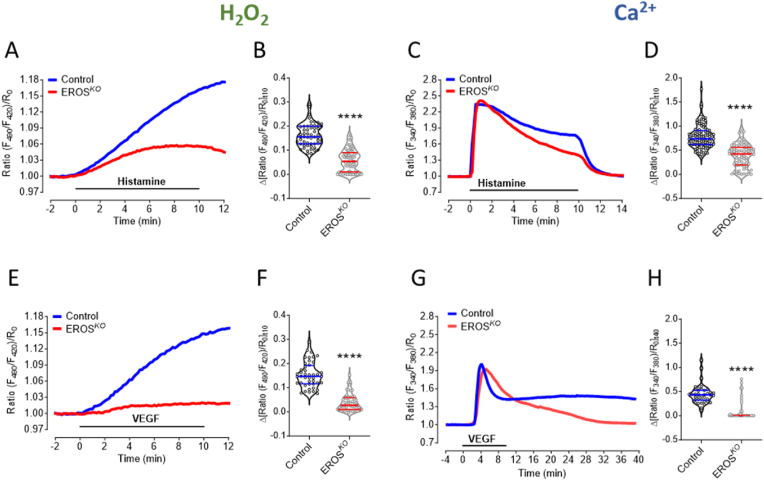
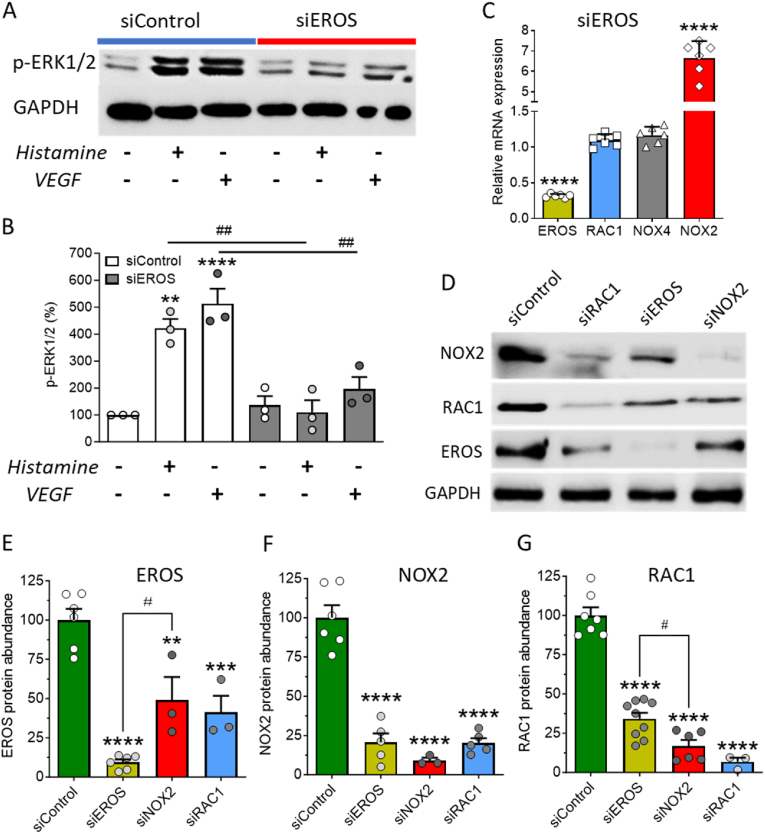
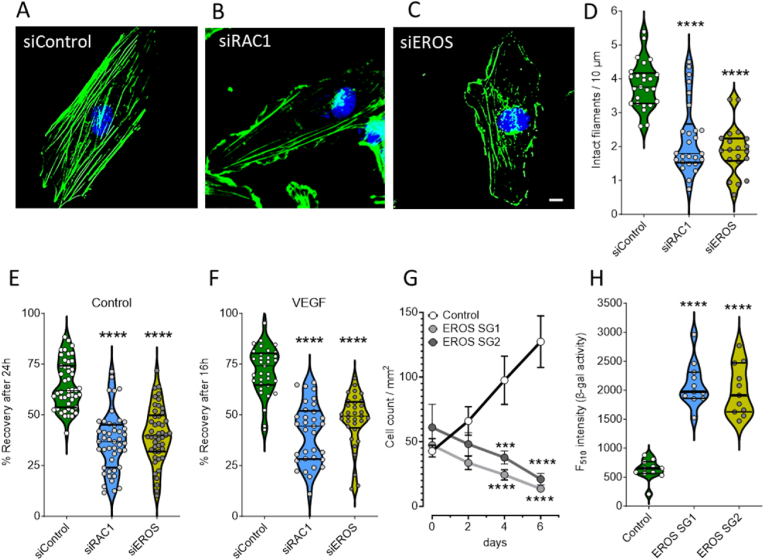
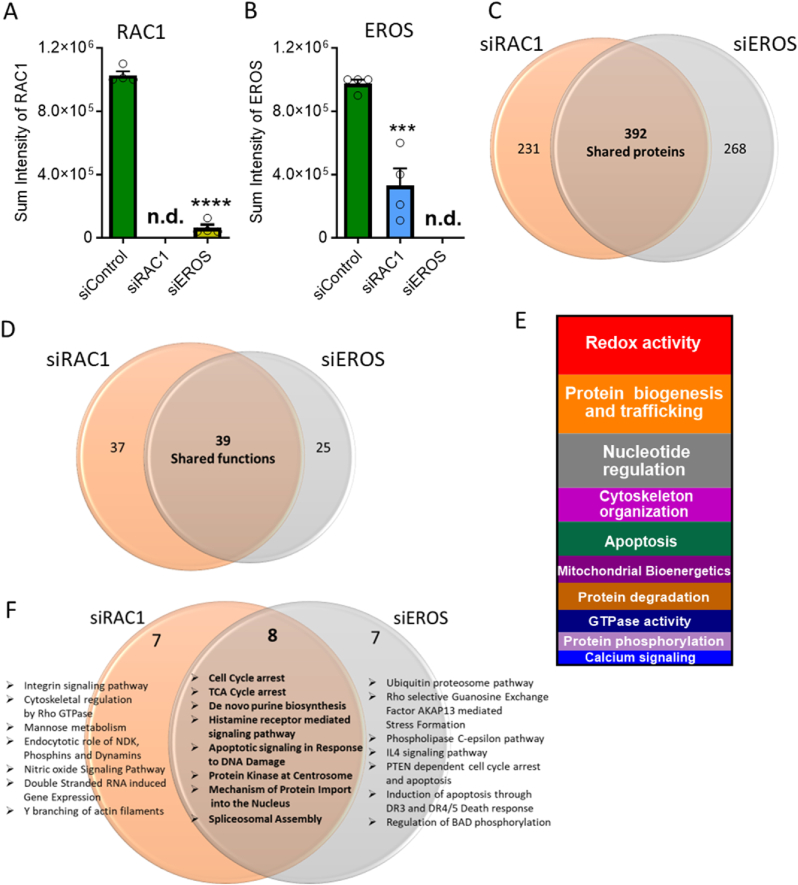
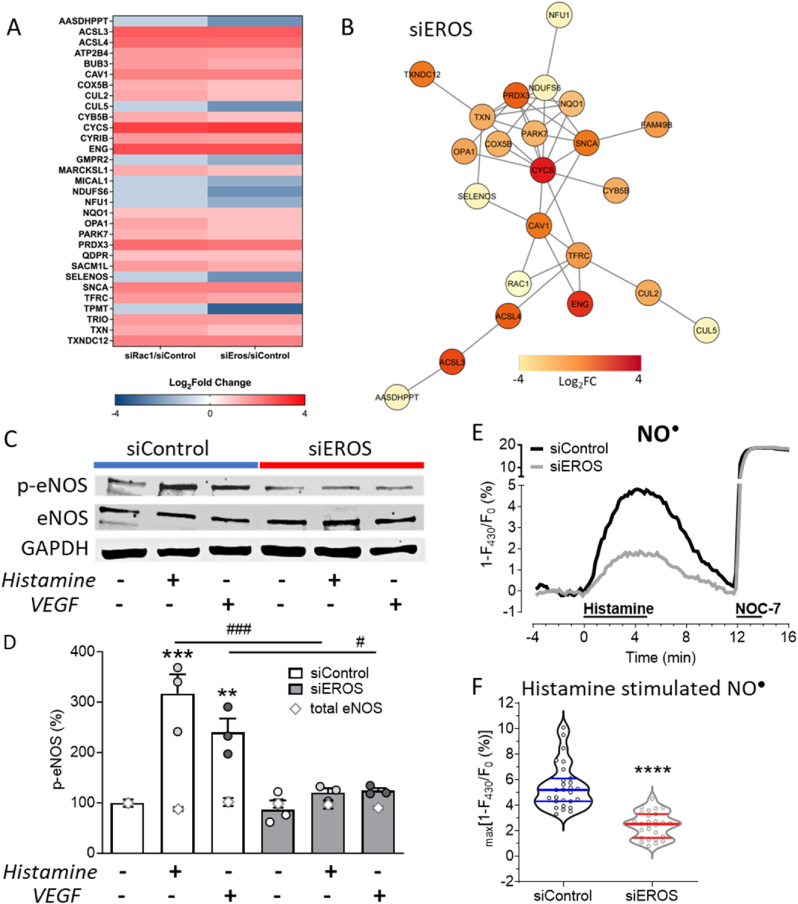
The chaperone protein EROS ("Essential for Reactive Oxygen Species") was recently discovered in phagocytes. EROS was shown to regulate the abundance of the ROS-producing enzyme NADPH oxidase isoform 2 (NOX2) and to control ROS-mediated cell killing. Reactive oxygen species are important not only in immune surveillance, but also modulate physiological signaling responses in multiple tissues. The roles of EROS have not been previously explored in the context of oxidant-modulated cell signaling. Here we show that EROS plays a key role in ROS-dependent signal transduction in vascular endothelial cells. We used siRNA-mediated knockdown and developed CRISPR/Cas9 knockout of EROS in human umbilical vein endothelial cells (HUVEC), both of which cause a significant decrease in the abundance of NOX2 protein, associated with a marked decrease in RAC1, a small G protein that activates NOX2. Loss of EROS also attenuates receptor-mediated hydrogen peroxide (H2O2) and Ca2+ signaling, disrupts cytoskeleton organization, decreases cell migration, and promotes cellular senescence. EROS knockdown blocks agonist-modulated eNOS phosphorylation and nitric oxide (NO●) generation. These effects of EROS knockdown are strikingly similar to the alterations in endothelial cell responses that we previously observed following RAC1 knockdown. Proteomic analyses following EROS or RAC1 knockdown in endothelial cells showed that reduced abundance of these two distinct proteins led to largely overlapping effects on endothelial biological processes, including oxidoreductase, protein phosphorylation, and endothelial nitric oxide synthase (eNOS) pathways. These studies demonstrate that EROS plays a central role in oxidant-modulated endothelial cell signaling by modulating NOX2 and RAC1.
Keywords: EROS; Endothelial cells; NOX2; RAC1; Redox signaling; Reductive stress.
Copyright © 2024 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare no competing financial interests.
Figures
References
-
- Thomas D.C., Clare S., Sowerby J.M., Pardo M., Juss J.K., Goulding D.A., van der Weyden L., Storisteanu D., Prakash A., Espéli M., Flint S., Lee J.C., Hoenderdos K., Kane L., Harcourt K., Mukhopadhyay S., Umrania Y., Antrobus R., Nathan J.A., Adams D.J., Bateman A., Choudhary J.S., Lyons P.A., Condliffe A.M., Chilvers E.R., Dougan G., Smith K.G.C. Eros is a novel transmembrane protein that controls the phagocyte respiratory burst and is essential for innate immunity. J. Exp. Med. 2017;214:1111–1128. doi: 10.1084/jem.20161382. - DOI - PMC - PubMed
-
- Randzavola L.O., Mortimer P.M., Garside E., Dufficy E.R., Schejtman A., Roumelioti G., Yu L., Pardo M., Spirohn K., Tolley C., Brandt C., Harcourt K., Nichols E., Nahorski M., Woods G., Williamson J.C., Suresh S., Sowerby J.M., Matsumoto M., Santos C.X.C., Kiar C.S., Mukhopadhyay S., Rae W.M., Dougan G.J., Grainger J., Lehner P.J., Calderwood M.A., Choudhary J., Clare S., Speak A., Santilli G., Bateman A., Smith K.G.C., Magnani F., Thomas D.C. EROS is a selective chaperone regulating the phagocyte NADPH oxidase and purinergic signalling. Elife. 2022;11:1–31. doi: 10.7554/eLife.76387. - DOI - PMC - PubMed
-
- Thomas D.C., Charbonnier L.M., Schejtman A., Aldhekri H., Coomber E.L., Dufficy E.R., Beenken A.E., Lee J.C., Clare S., Speak A.O., Thrasher A.J., Santilli G., Al-Mousa H., Alkuraya F.S., Chatila T.A., Smith K.G.C. EROS/CYBC1 mutations: decreased NADPH oxidase function and chronic granulomatous disease. J. Allergy Clin. Immunol. 2019;143:782–785.e1. doi: 10.1016/j.jaci.2018.09.019. - DOI - PMC - PubMed
-
- Arnadottir G.A., Norddahl G.L., Gudmundsdottir S., Agustsdottir A.B., Sigurdsson S., Jensson B.O., Bjarnadottir K., Theodors F., Benonisdottir S., Ivarsdottir E.V., Oddsson A., Kristjansson R.P., Sulem G., Alexandersson K.F., Juliusdottir T., Gudmundsson K.R., Saemundsdottir J., Jonasdottir A., Jonasdottir A., Sigurdsson A., Manzanillo P., Gudjonsson S.A., Thorisson G.A., Magnusson O.T., Masson G., Orvar K.B., Holm H., Bjornsson S., Arngrimsson R., Gudbjartsson D.F., Thorsteinsdottir U., Jonsdottir I., Haraldsson A., Sulem P., Stefansson K. A homozygous loss-of-function mutation leading to CYBC1 deficiency causes chronic granulomatous disease. Nat. Commun. 2018;9:1–9. doi: 10.1038/s41467-018-06964-x. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
