

Multiscale selection in spatially structured populations
- PMID: 38808450
- PMCID: PMC11285734
- DOI: 10.1098/rspb.2023.2559
Multiscale selection in spatially structured populations
Abstract
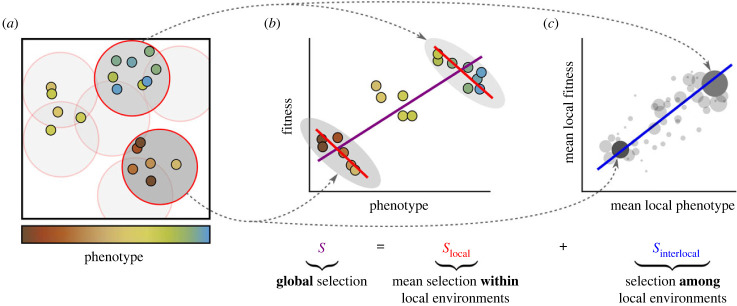
The spatial structure of populations is key to many (eco-)evolutionary processes. In such cases, the strength and sign of selection on a trait may depend on the spatial scale considered. An example is the evolution of altruism: selection in local environments often favours cheaters over altruists, but this can be outweighed by selection at larger scales, favouring clusters of altruists over clusters of cheaters. For populations subdivided into distinct groups, this effect is described formally by multilevel selection theory. However, many populations do not consist of non-overlapping groups but rather (self-)organize into other ecological patterns. We therefore present a mathematical framework for multiscale selection. This framework decomposes natural selection into two parts: local selection, acting within environments of a certain size, and interlocal selection, acting among them. Varying the size of the local environments subsequently allows one to measure the contribution to selection of each spatial scale. To illustrate the use of this framework, we apply it to models of the evolution of altruism and pathogen transmissibility. The analysis identifies how and to what extent ecological processes at different spatial scales contribute to selection and compete, thus providing a rigorous underpinning to eco-evolutionary intuitions.
Keywords: Price’s equation; altruism; evolution; pathogen transmissibility; self-organization; spatial structure.
Conflict of interest statement
We declare we have no competing interests.
Figures


References
MeSH terms
LinkOut - more resources
Full Text Sources