

The phage shock protein (PSP) envelope stress response: discovery of novel partners and evolutionary history
- PMID: 38809013
- PMCID: PMC11237479
- DOI: 10.1128/msystems.00847-23
The phage shock protein (PSP) envelope stress response: discovery of novel partners and evolutionary history
Abstract
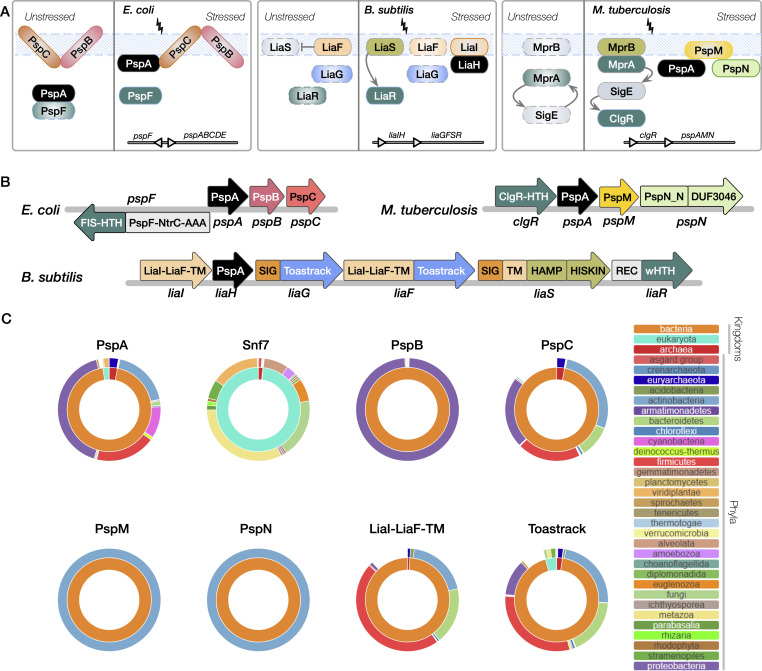
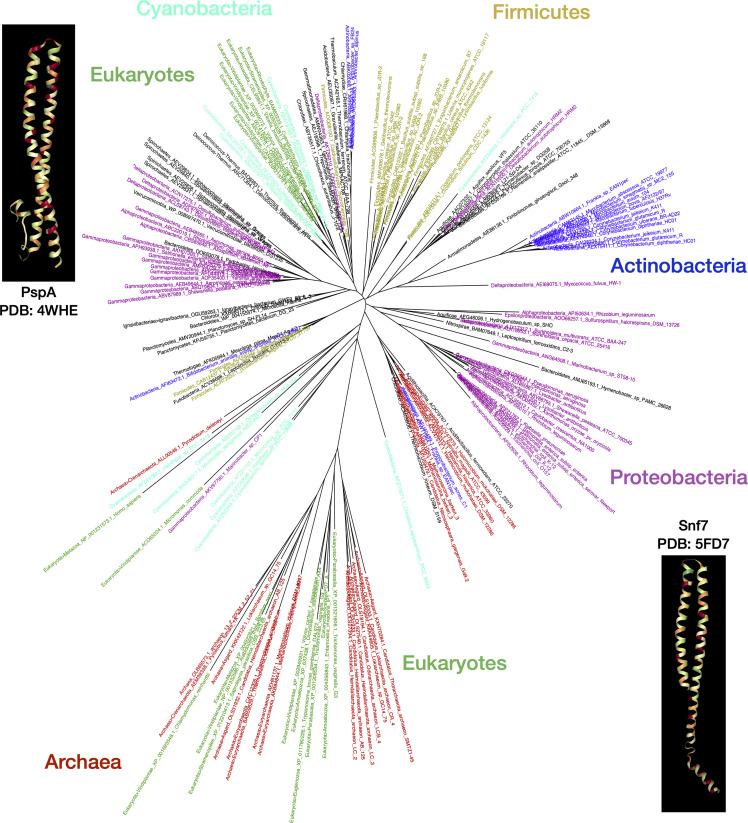
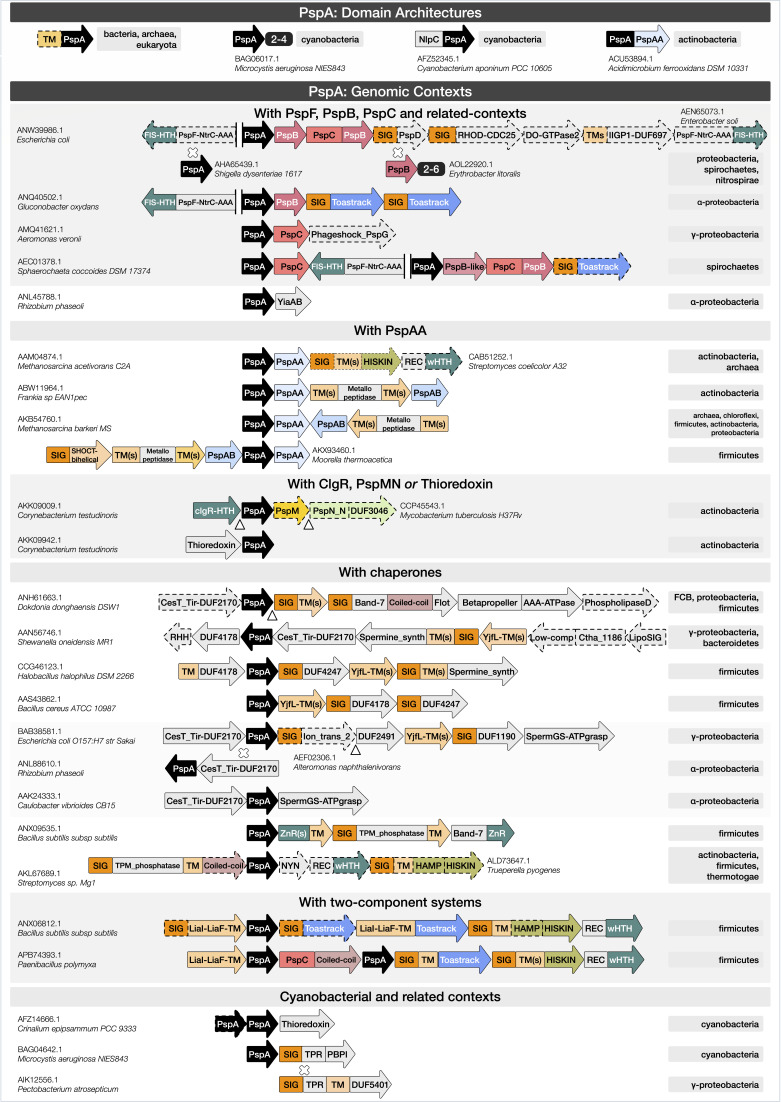
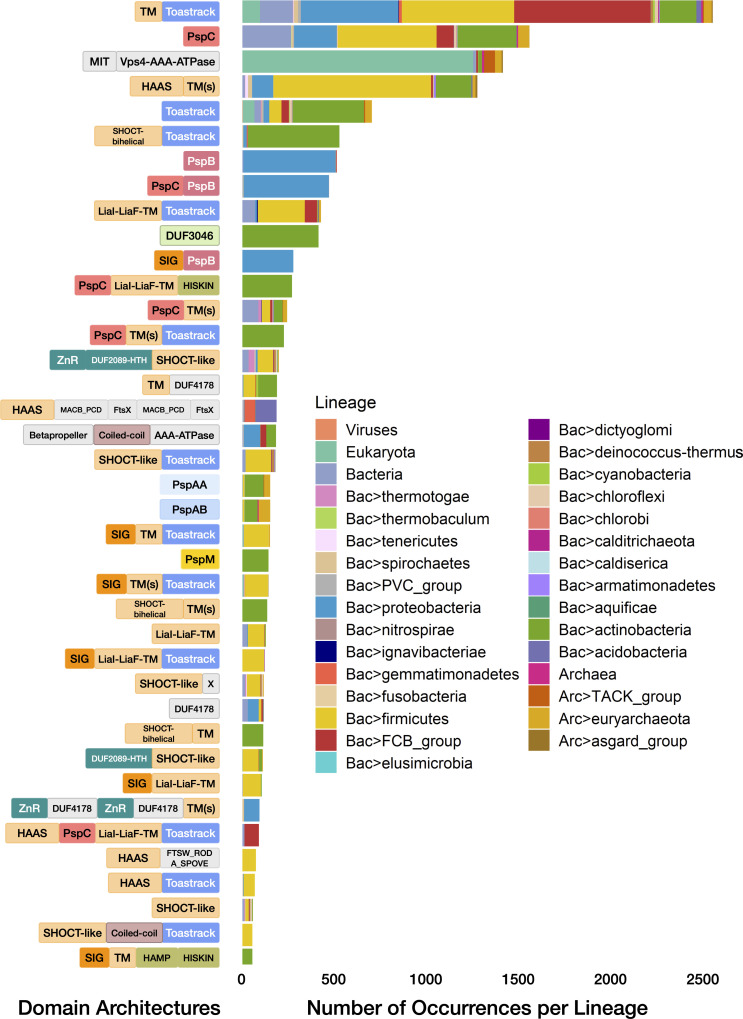
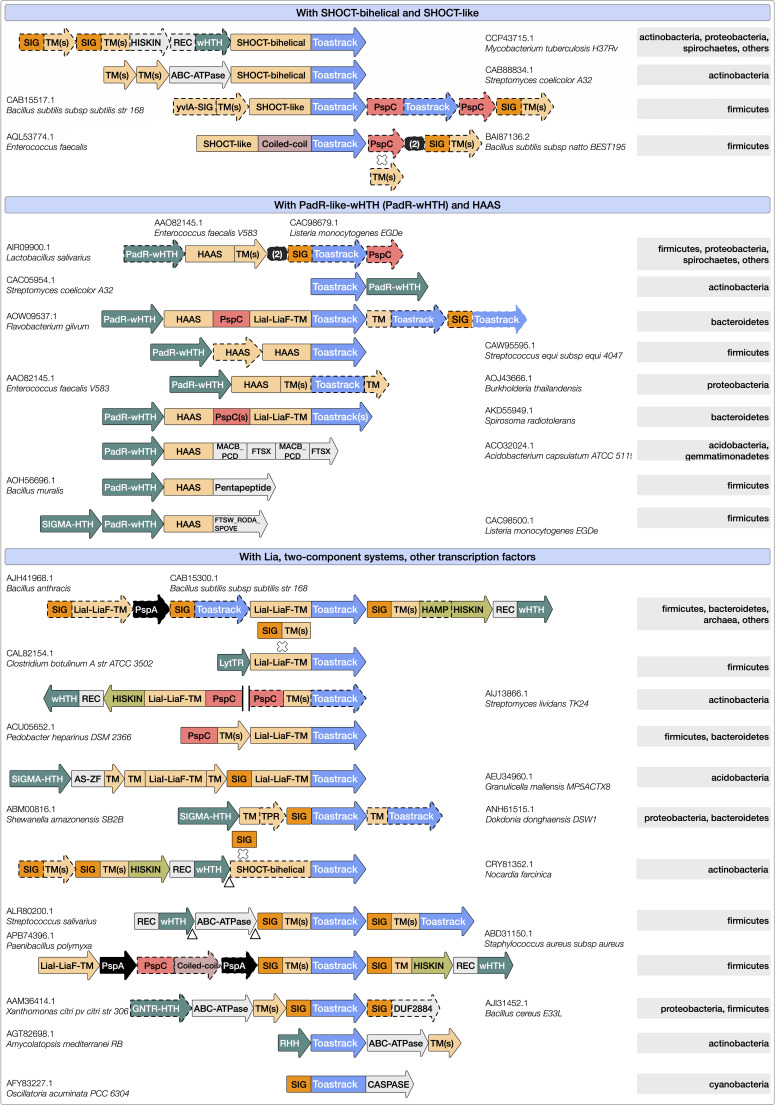
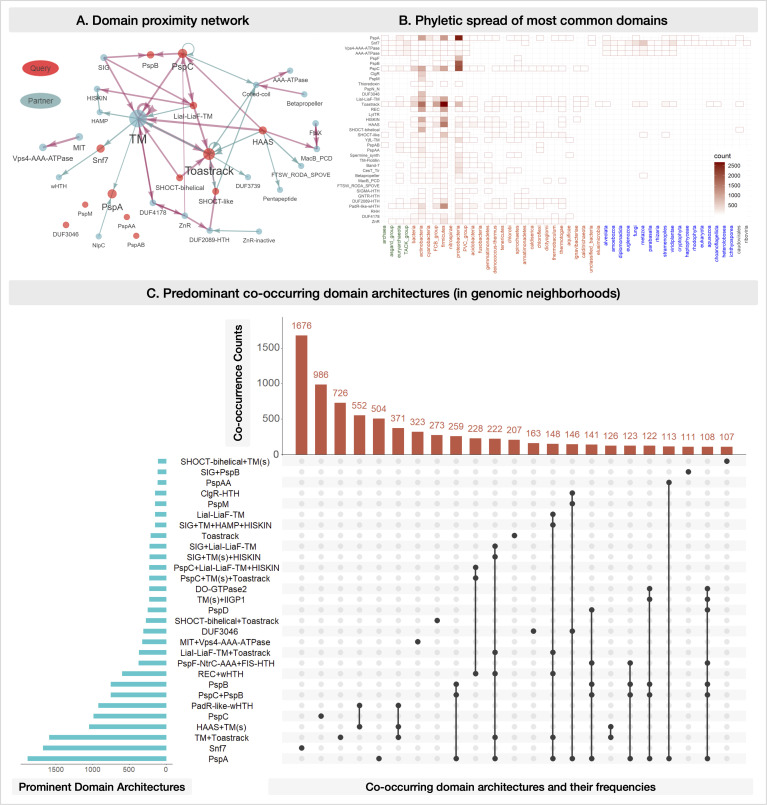
Bacterial phage shock protein (PSP) systems stabilize the bacterial cell membrane and protect against envelope stress. These systems have been associated with virulence, but despite their critical roles, PSP components are not well characterized outside proteobacteria. Using comparative genomics and protein sequence-structure-function analyses, we systematically identified and analyzed PSP homologs, phyletic patterns, domain architectures, and gene neighborhoods. This approach underscored the evolutionary significance of the system, revealing that its core protein PspA (Snf7 in ESCRT outside bacteria) was present in the last universal common ancestor and that this ancestral functionality has since diversified into multiple novel, distinct PSP systems across life. Several novel partners of the PSP system were identified: (i) the Toastrack domain, likely facilitating assembly of sub-membrane stress-sensing and signaling complexes, (ii) the newly defined HTH-associated α-helical signaling domain-PadR-like transcriptional regulator pair system, and (iii) multiple independent associations with ATPase, CesT/Tir-like chaperone, and Band-7 domains in proteins thought to mediate sub-membrane dynamics. Our work also uncovered links between the PSP components and other domains, such as novel variants of SHOCT-like domains, suggesting roles in assembling membrane-associated complexes of proteins with disparate biochemical functions. Results are available at our interactive web app, https://jravilab.org/psp.IMPORTANCEPhage shock proteins (PSP) are virulence-associated, cell membrane stress-protective systems. They have mostly been characterized in Proteobacteria and Firmicutes. We now show that a minimal PSP system was present in the last universal common ancestor that evolved and diversified into newly identified functional contexts. Recognizing the conservation and evolution of PSP systems across bacterial phyla contributes to our understanding of stress response mechanisms in prokaryotes. Moreover, the newly discovered PSP modularity will likely prompt new studies of lineage-specific cell envelope structures, lifestyles, and adaptation mechanisms. Finally, our results validate the use of domain architecture and genetic context for discovery in comparative genomics.
Keywords: PSP; comparative genomics; domain architectures; envelope stress response; genomic contexts and neighborhoods; molecular evolution; phage shock protein response; phylogeny.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Variations on a theme: evolution of the phage-shock-protein system in Actinobacteria.Antonie Van Leeuwenhoek. 2018 May;111(5):753-760. doi: 10.1007/s10482-018-1053-5. Epub 2018 Feb 27. Antonie Van Leeuwenhoek. 2018. PMID: 29488183 Free PMC article.
-
Short-Term Memory Impairment.2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2024 Jun 8. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31424720 Free Books & Documents.
-
The Black Book of Psychotropic Dosing and Monitoring.Psychopharmacol Bull. 2024 Jul 8;54(3):8-59. Psychopharmacol Bull. 2024. PMID: 38993656 Free PMC article. Review.
-
Systemic Inflammatory Response Syndrome.2025 Jun 20. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. 2025 Jun 20. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2025 Jan–. PMID: 31613449 Free Books & Documents.
-
Single-incision sling operations for urinary incontinence in women.Cochrane Database Syst Rev. 2017 Jul 26;7(7):CD008709. doi: 10.1002/14651858.CD008709.pub3. Cochrane Database Syst Rev. 2017. Update in: Cochrane Database Syst Rev. 2023 Oct 27;10:CD008709. doi: 10.1002/14651858.CD008709.pub4. PMID: 28746980 Free PMC article. Updated.
Cited by
-
BacA: a possible regulator that contributes to the biofilm formation of Pseudomonas aeruginosa.Front Microbiol. 2024 Mar 5;15:1332448. doi: 10.3389/fmicb.2024.1332448. eCollection 2024. Front Microbiol. 2024. PMID: 38505547 Free PMC article.
-
An Stomatin, Prohibitin, Flotillin, and HflK/C-Domain Protein Required to Link the Phage-Shock Protein to the Membrane in Bacillus subtilis.Front Microbiol. 2021 Oct 28;12:754924. doi: 10.3389/fmicb.2021.754924. eCollection 2021. Front Microbiol. 2021. PMID: 34777311 Free PMC article.
-
Identification of Clostridioides difficile mutants with increased daptomycin resistance.J Bacteriol. 2024 Mar 21;206(3):e0036823. doi: 10.1128/jb.00368-23. Epub 2024 Feb 20. J Bacteriol. 2024. PMID: 38376203 Free PMC article.
-
Functional amyloid proteins confer defence against predatory bacteria.Nature. 2025 Aug;644(8075):197-204. doi: 10.1038/s41586-025-09204-7. Epub 2025 Jul 2. Nature. 2025. PMID: 40604283
-
Engineered Phage Enables Efficient Control of Gene Expression upon Infection of the Host Cell.Int J Mol Sci. 2024 Dec 30;26(1):250. doi: 10.3390/ijms26010250. Int J Mol Sci. 2024. PMID: 39796105 Free PMC article.
References
MeSH terms
Substances
Grants and funding
- R01AI104615/HHS | National Institutes of Health (NIH)
- R01 HL106788/HL/NHLBI NIH HHS/United States
- Start-up funds/University of Colorado Anschutz (CU)
- R01 HL149450/HL/NHLBI NIH HHS/United States
- R01HL106788/HHS | National Institutes of Health (NIH)
- R01 AI104615/AI/NIAID NIH HHS/United States
- R01HL149450/HHS | National Institutes of Health (NIH)
- Oak Ridge Institute for Science and Education Scholarship/Oak Ridge Associated Universities (ORAU)
- Intramural Research Program/HHS | National Institutes of Health (NIH)
- College of Veterinary Medicine Endowed Research Funds/Michigan State University (MSU)
LinkOut - more resources
Full Text Sources
Miscellaneous
