

TGF-β signalling limits effector function capacity of NK cell anti-tumour immunity in human bladder cancer
- PMID: 38810560
- PMCID: PMC11154198
- DOI: 10.1016/j.ebiom.2024.105176
TGF-β signalling limits effector function capacity of NK cell anti-tumour immunity in human bladder cancer
Abstract
Background: Natural killer (NK) cells are important innate immunity players and have unique abilities to recognize and eliminate cancer cells, particularly in settings of antibody-opsonization and antibody-dependant cellular cytotoxicity (ADCC). However, NK cell-based responses in bladder cancers to therapeutic antibodies are typically immunosuppressed, and these immunosuppressive mechanisms are largely unknown.
Methods: Single cell RNA sequencing (scRNA-seq) and high-dimensional flow cytometry were used to investigate the phenotype of tumour-infiltrating NK cells in patients with bladder cancer. Further, in vitro, and in vivo models of this disease were used to validate these findings.
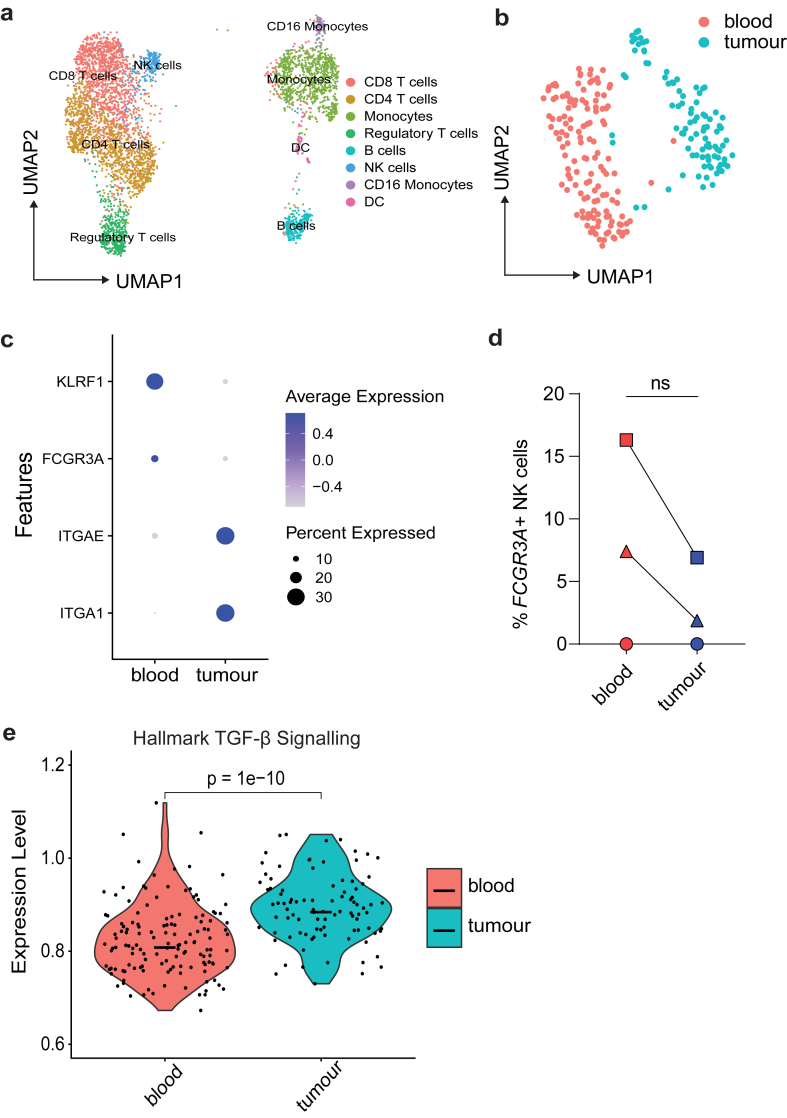
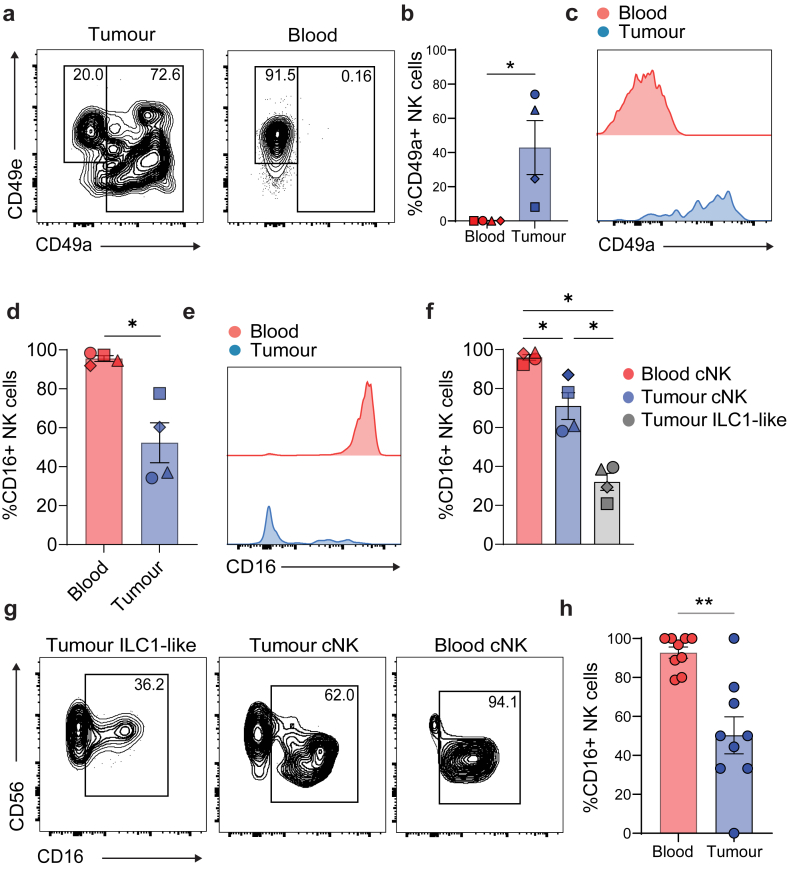
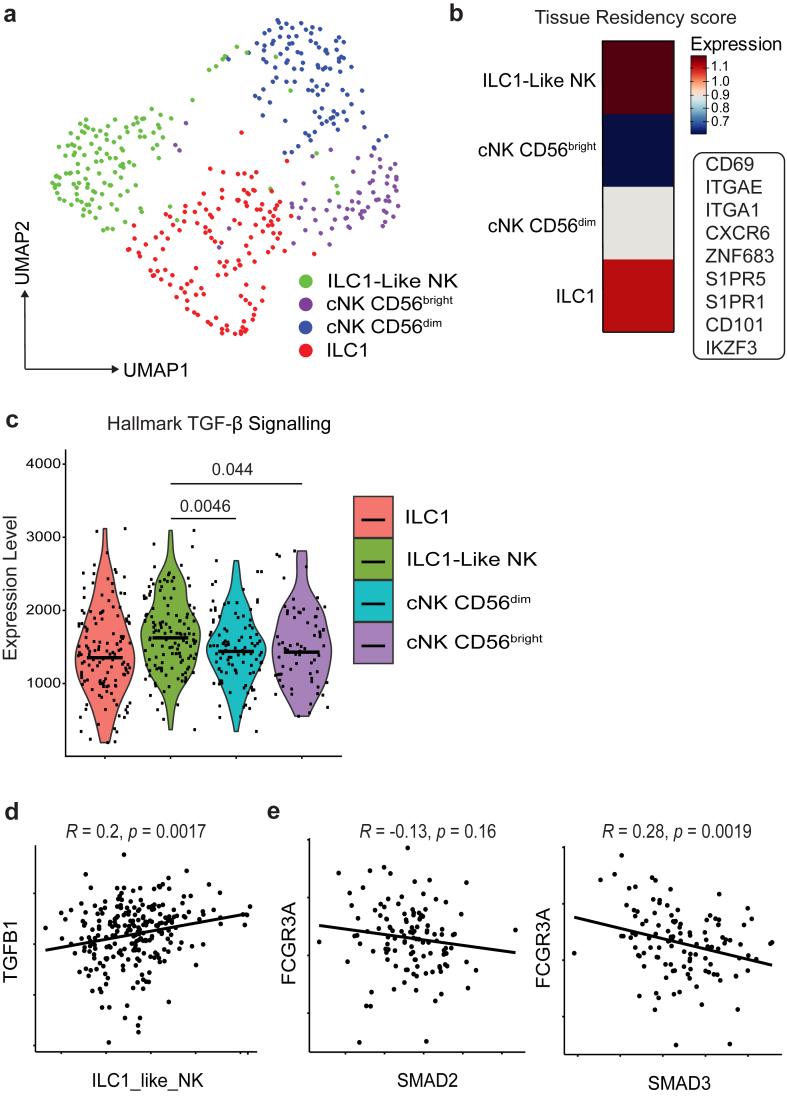
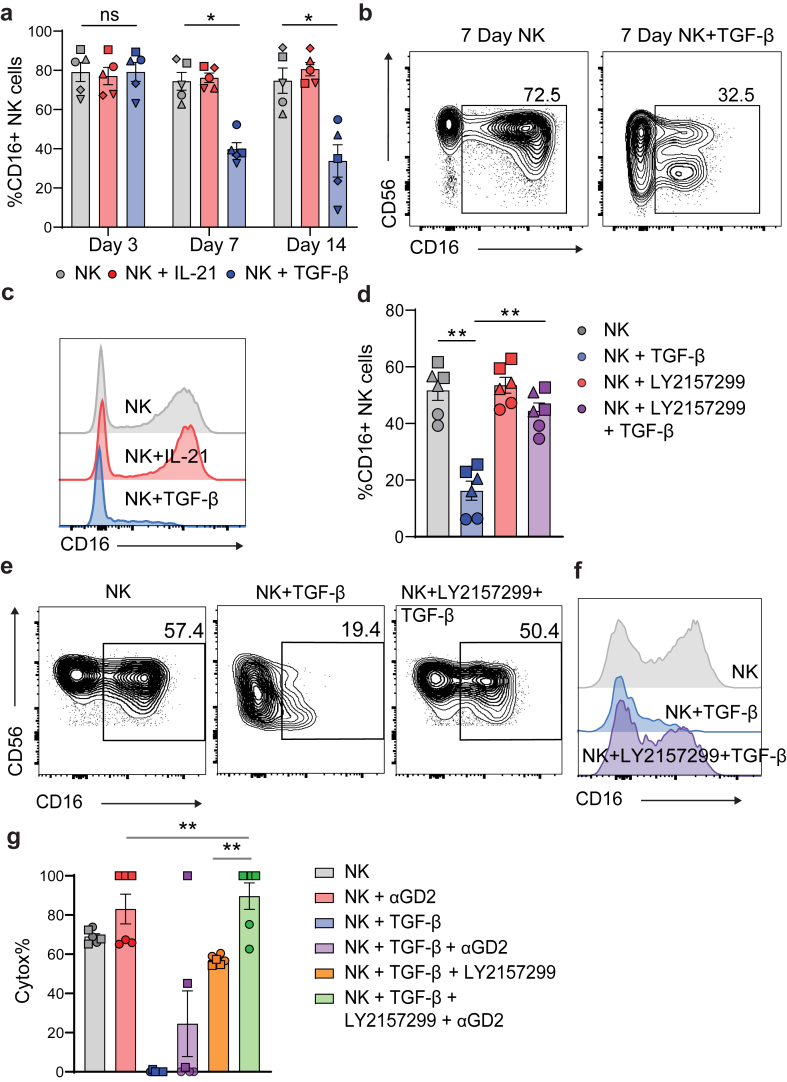
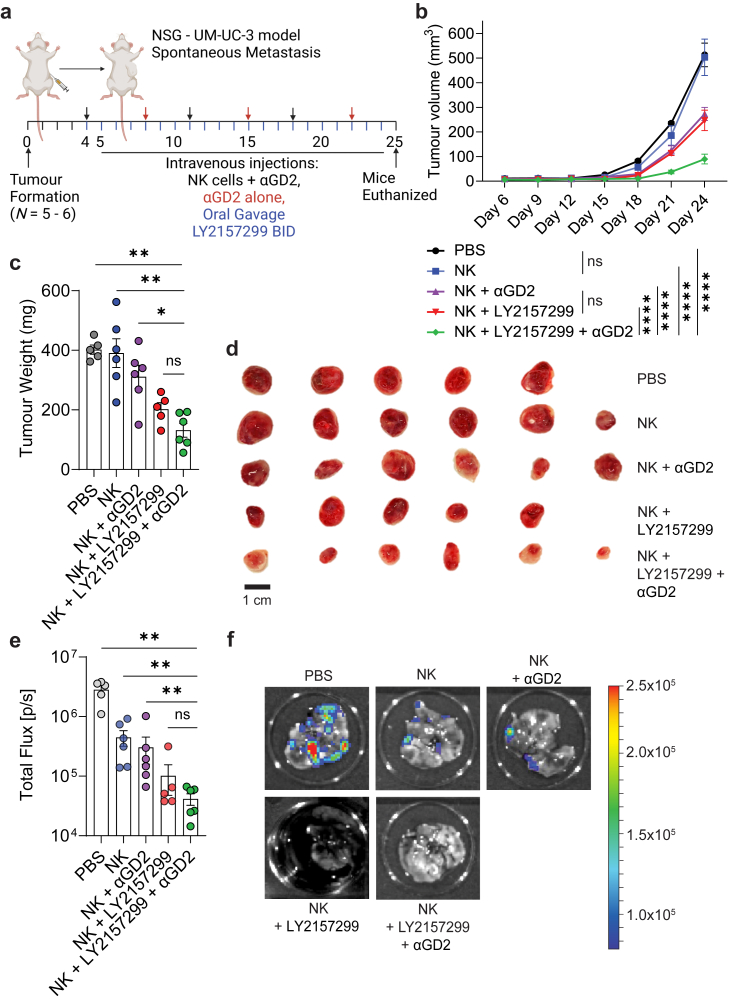
Findings: NK cells within bladder tumours displayed reduced expression of FcγRIIIa/CD16, the critical Fc receptor involved in ADCC-mediated cytotoxicity, on both transcriptional and protein levels. Transcriptional signatures of transforming growth factor (TGF)-β-signalling, a pleiotropic cytokine known for its immunosuppressive and tissue residency-inducing effects, were upregulated in tumour-infiltrating NK cells. TGF-β mediated CD16 downregulation on NK cells, was further validated in vitro, which was accompanied by a transition into a tissue residency phenotype. This CD16 downregulation was also abrogated by TGF-βR signalling inhibition, which could also restore the ADCC ability of NK cells subject to TGF-β effects. In a humanized mouse model of bladder cancer, mice treated with a TGF-β inhibitor exhibited increased ADCC activity compared to mice treated only with antibodies.
Interpretation: This study highlights how TGF-β-rich bladder cancers inhibit NK cell-mediated ADCC by downregulating CD16. TGF-β inhibition represents new avenues to reverse immunosuppression and enhance the tumoricidal capacity of NK cells in bladder cancer.
Funding: The Guimaraes Laboratory is funded by a US Department of Defense-Breast Cancer Research Program-Breakthrough Award Level 1 (#BC200025), a grant (#2019485) awarded through the Medical Research Future Fund (MRFF, with the support of the Queensland Children's Hospital Foundation, Microba Life Sciences, Richie's Rainbow Foundation, Translational Research Institute (TRI) and UQ), and a grant (#RSS_2023_085) funded by a Metro South Health Research Support Scheme. J.K.M.W. is funded by a UQ Research Training Program PhD Scholarship and N.O. is funded by a NHMRC Postgraduate Scholarship (#2021932).
Keywords: ADCC; Bladder cancer; CD16; NK cells; TGF-β.
Copyright © 2024 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of interests A/Prof. F. Simpson declares support from NHMRC Investigator grant (#2026628) and holds patents unrelated to this manuscript. She serves on advisory boards (Elsevier, UQ Academic Board, Victorian Cancer Council and NSW Cancer Council); and none of these roles' present conflicts with this study. A/Prof. Guimaraes' laboratory is funded by multiple grants, including the US Department of Defense and Medical Research Future Fund. He consults for Microba Life Sciences and Prescient Therapeutics, with all payments made to his institution and unrelated to this study. Both authors confirm that these disclosures do not influence the findings of this manuscript. The remaining authors declared that there is no conflict of interest related to this work.
Figures
References
-
- Souza-Fonseca-Guimaraes F., Cursons J., Huntington N.D. The emergence of natural killer cells as a major target in cancer immunotherapy. Trends Immunol. 2019;40(2):142–158. - PubMed
-
- Wong J.K.M., Dolcetti R., Rhee H., Simpson F., Souza-Fonseca-Guimaraes F. Weaponizing natural killer cells for solid cancer immunotherapy. Trends in Cancer. 2023;9(2):111–121. - PubMed
-
- Vivier E., Rebuffet L., Narni-Mancinelli E., Cornen S., Igarashi R.Y., Fantin V.R. Natural killer cell therapies. Nature. 2024;626(8000):727–736. - PubMed
-
- Eckl J., Buchner A., Prinz P.U., et al. Transcript signature predicts tissue NK cell content and defines renal cell carcinoma subgroups independent of TNM staging. J Mol Med. 2012;90(1):55–66. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
