

Evolution of diapause in the African turquoise killifish by remodeling the ancient gene regulatory landscape
- PMID: 38810644
- PMCID: PMC11970524
- DOI: 10.1016/j.cell.2024.04.048
Evolution of diapause in the African turquoise killifish by remodeling the ancient gene regulatory landscape
Abstract
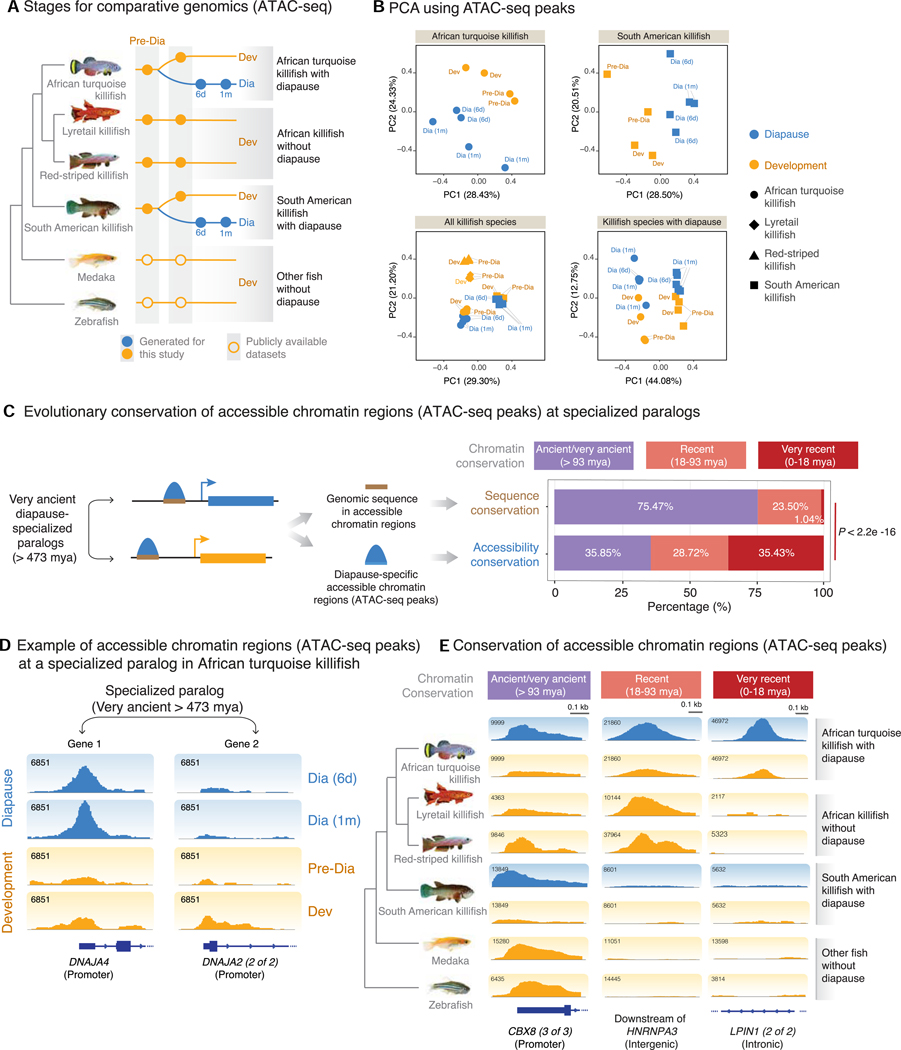
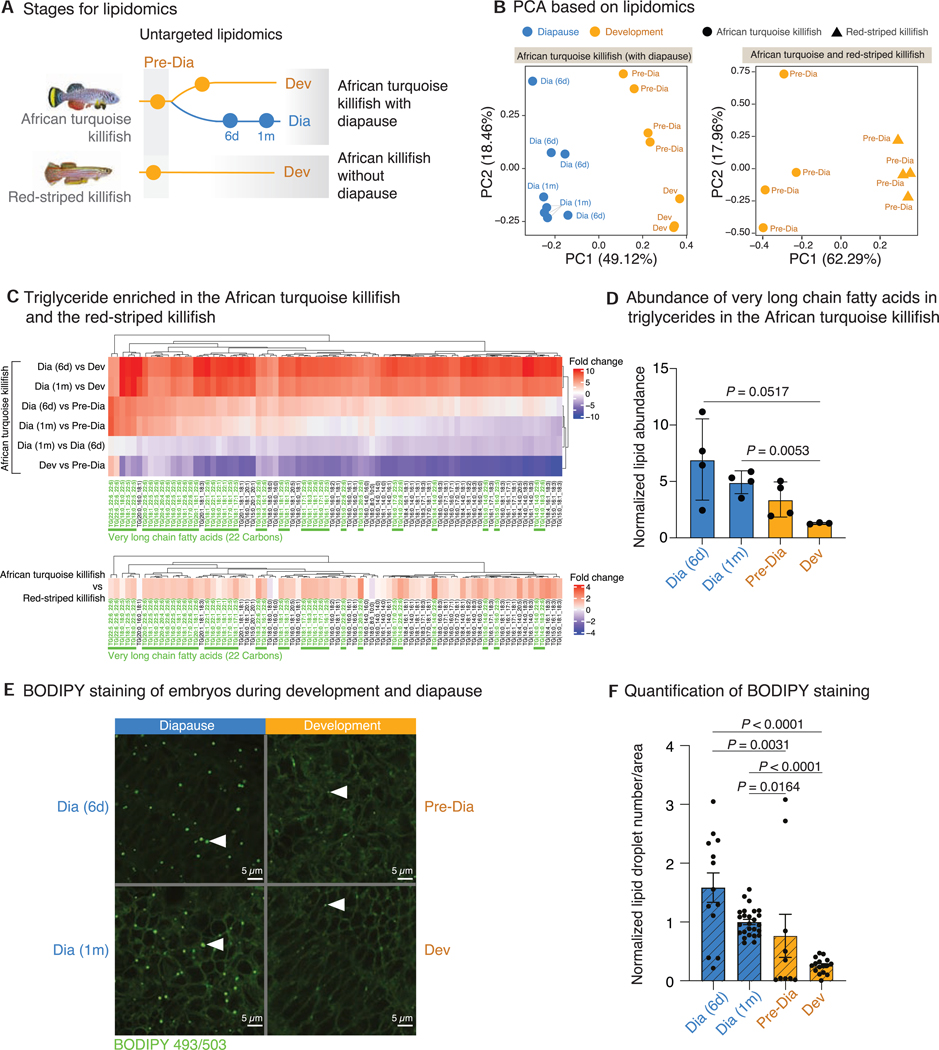
Suspended animation states allow organisms to survive extreme environments. The African turquoise killifish has evolved diapause as a form of suspended development to survive a complete drought. However, the mechanisms underlying the evolution of extreme survival states are unknown. To understand diapause evolution, we performed integrative multi-omics (gene expression, chromatin accessibility, and lipidomics) in the embryos of multiple killifish species. We find that diapause evolved by a recent remodeling of regulatory elements at very ancient gene duplicates (paralogs) present in all vertebrates. CRISPR-Cas9-based perturbations identify the transcription factors REST/NRSF and FOXOs as critical for the diapause gene expression program, including genes involved in lipid metabolism. Indeed, diapause shows a distinct lipid profile, with an increase in triglycerides with very-long-chain fatty acids. Our work suggests a mechanism for the evolution of complex adaptations and offers strategies to promote long-term survival by activating suspended animation programs in other species.
Keywords: aging; comparative genomics; complex trait evolution; diapause; epigenetics; epigenomics; killifish; lipidomics; metabolomics; suspended animation.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.B. is a scientific advisory board member of Calico. M.P.S. is a co-founder and the scientific advisory board member of Personalis, Qbio, January AI, SensOmics, Filtricine, Protos, Mirvie, Onza, Marble Therapeutics, Iollo, and NextThought AI. He is also on the scientific advisory board of Jupiter, Applied Cognition, Neuvivo, Mitrix, and Enovone. K.C. is currently an AstraZeneca employee.
Figures





Similar articles
-
Analysis of microRNA expression reveals convergent evolution of the molecular control of diapause in annual killifishes.Front Genet. 2025 Jul 3;16:1583989. doi: 10.3389/fgene.2025.1583989. eCollection 2025. Front Genet. 2025. PMID: 40678384 Free PMC article.
-
Vertebrate diapause preserves organisms long term through Polycomb complex members.Science. 2020 Feb 21;367(6480):870-874. doi: 10.1126/science.aaw2601. Science. 2020. PMID: 32079766 Free PMC article.
-
Analysis of Different Strains of the Turquoise Killifish Identify Transcriptomic Signatures Associated With Heritable Lifespan Differences.J Gerontol A Biol Sci Med Sci. 2025 Jun 10;80(7):glae255. doi: 10.1093/gerona/glae255. J Gerontol A Biol Sci Med Sci. 2025. PMID: 39780413
-
The African turquoise killifish (Nothobranchius furzeri): A model of age-related diseases and declining regenerative capability across multiple organs.Mech Ageing Dev. 2025 Aug;226:112087. doi: 10.1016/j.mad.2025.112087. Epub 2025 Jun 30. Mech Ageing Dev. 2025. PMID: 40602620 Review.
-
Systemic pharmacological treatments for chronic plaque psoriasis: a network meta-analysis.Cochrane Database Syst Rev. 2017 Dec 22;12(12):CD011535. doi: 10.1002/14651858.CD011535.pub2. Cochrane Database Syst Rev. 2017. Update in: Cochrane Database Syst Rev. 2020 Jan 9;1:CD011535. doi: 10.1002/14651858.CD011535.pub3. PMID: 29271481 Free PMC article. Updated.
Cited by
-
A conserved switch to less catalytically active Polycomb repressive complexes in non-dividing cells.Cell Rep. 2025 Jan 28;44(1):115192. doi: 10.1016/j.celrep.2024.115192. Epub 2025 Jan 11. Cell Rep. 2025. PMID: 39799569
-
Rapid and precise genome engineering in a naturally short-lived vertebrate.Elife. 2023 May 9;12:e80639. doi: 10.7554/eLife.80639. Elife. 2023. PMID: 37191291 Free PMC article.
-
The African Turquoise Killifish: A Scalable Vertebrate Model for Aging and Other Complex Phenotypes.Cold Spring Harb Protoc. 2024 Mar 1;2024(3):107737. doi: 10.1101/pdb.over107737. Cold Spring Harb Protoc. 2024. PMID: 37100468 Free PMC article.
-
FOXO1-mediated lipid metabolism maintains mammalian embryos in dormancy.Nat Cell Biol. 2024 Feb;26(2):181-193. doi: 10.1038/s41556-023-01325-3. Epub 2024 Jan 4. Nat Cell Biol. 2024. PMID: 38177284 Free PMC article.
-
Metabolic regulation in adult and aging skeletal muscle stem cells.Genes Dev. 2025 Feb 3;39(3-4):186-208. doi: 10.1101/gad.352277.124. Genes Dev. 2025. PMID: 39662967 Free PMC article. Review.
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
