

Tissue-specific landscape of protein aggregation and quality control in an aging vertebrate
- PMID: 38810654
- PMCID: PMC11265985
- DOI: 10.1016/j.devcel.2024.04.014
Tissue-specific landscape of protein aggregation and quality control in an aging vertebrate
Abstract
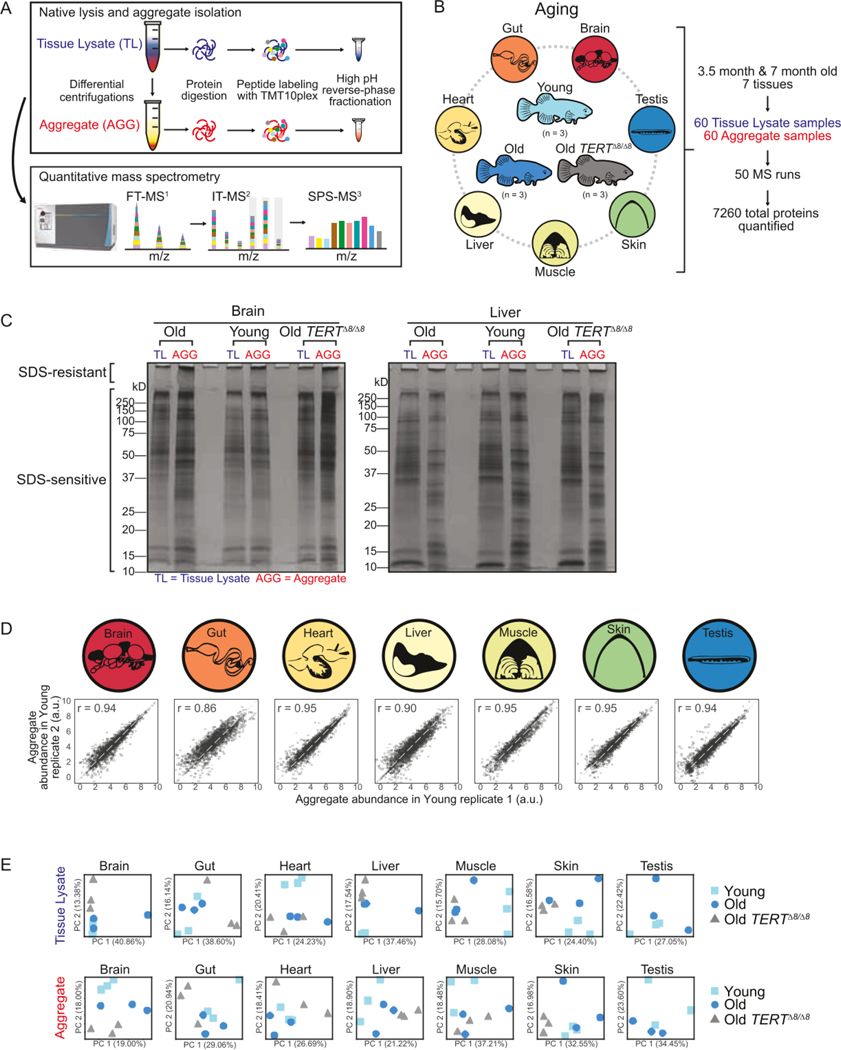
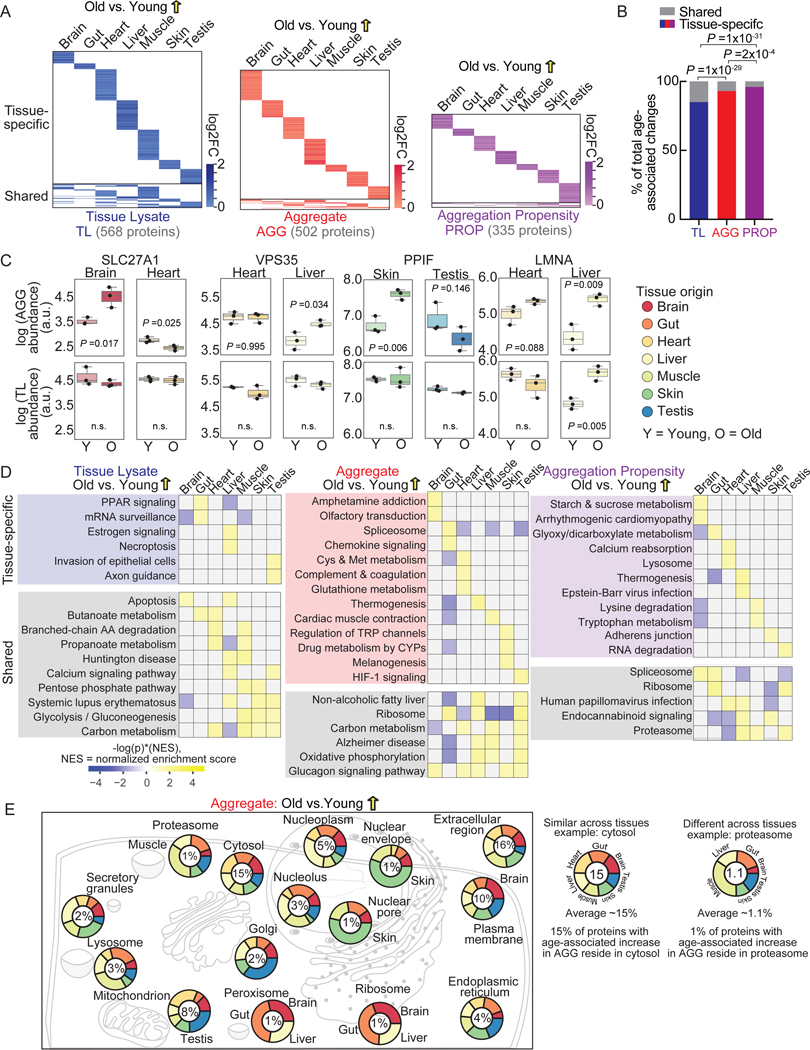
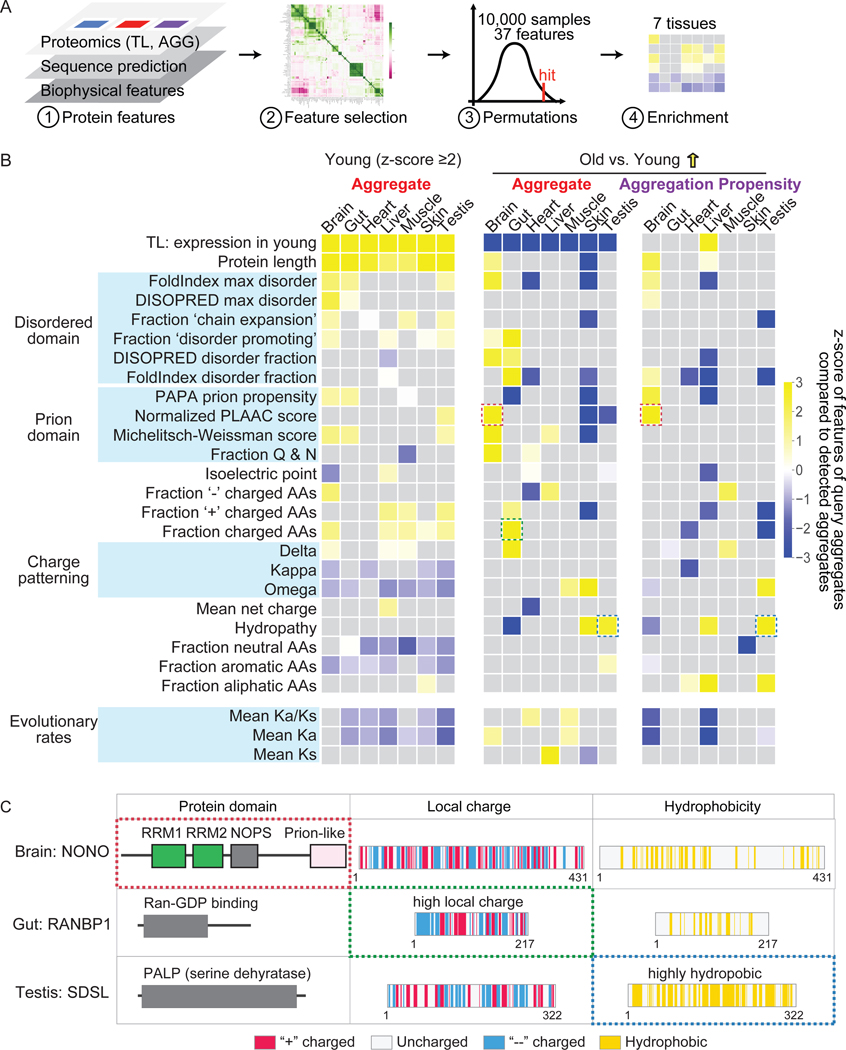
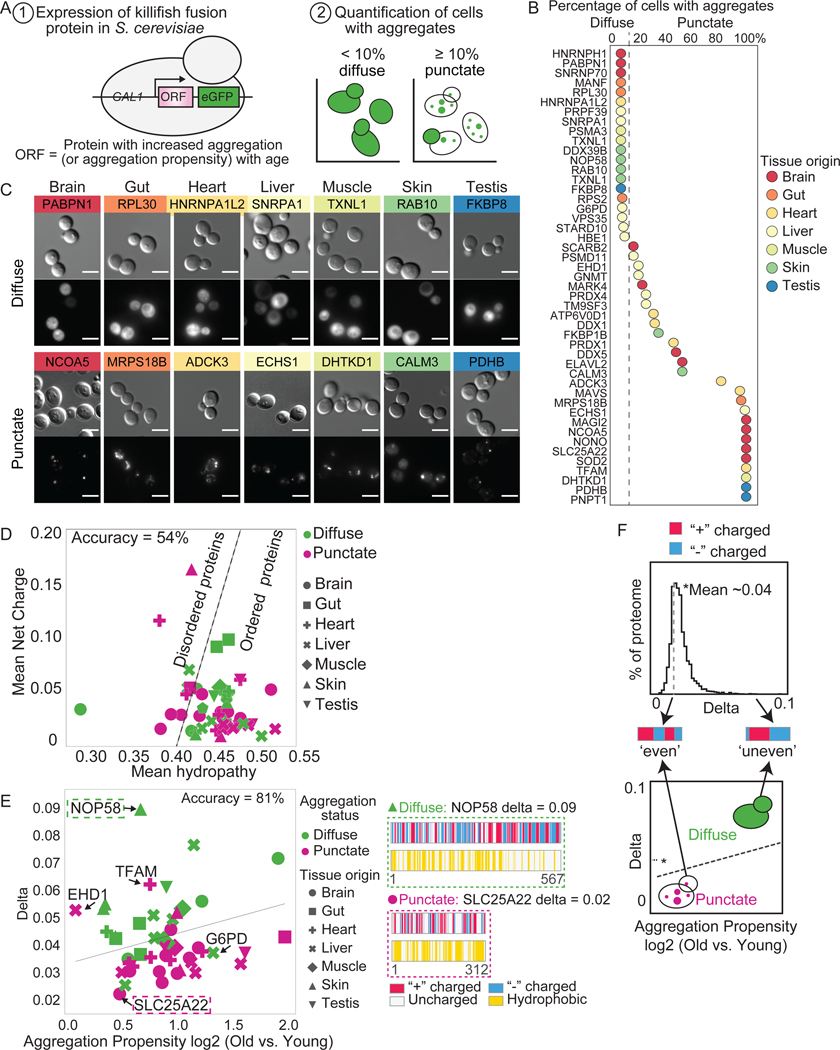
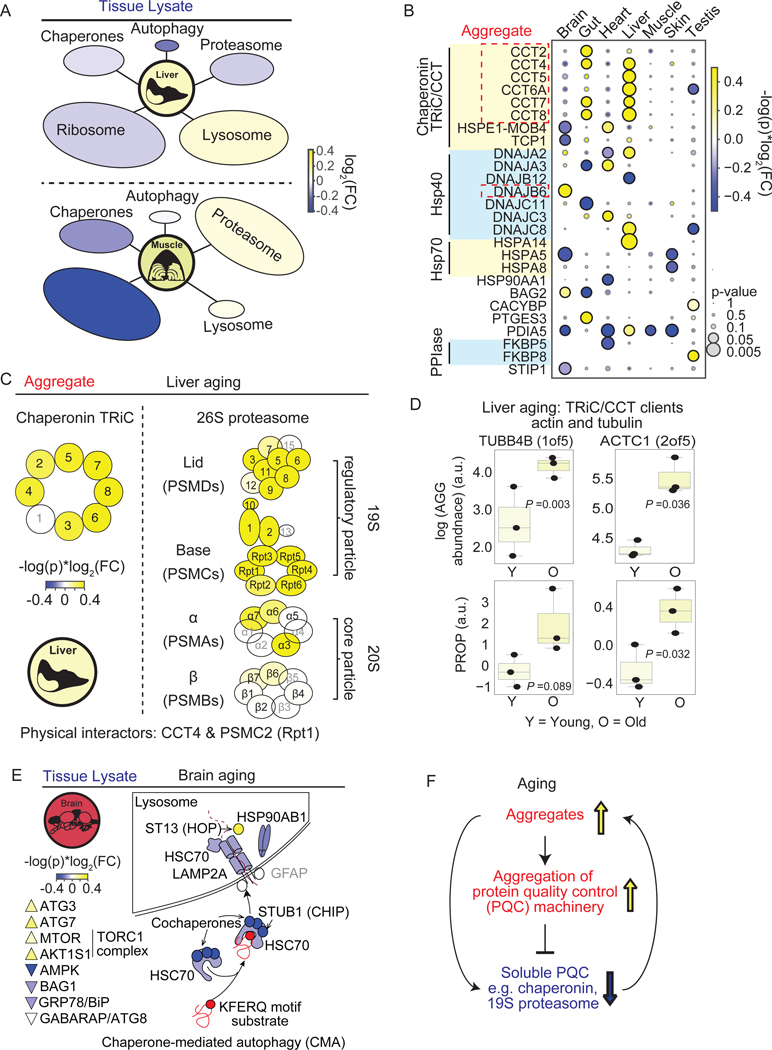
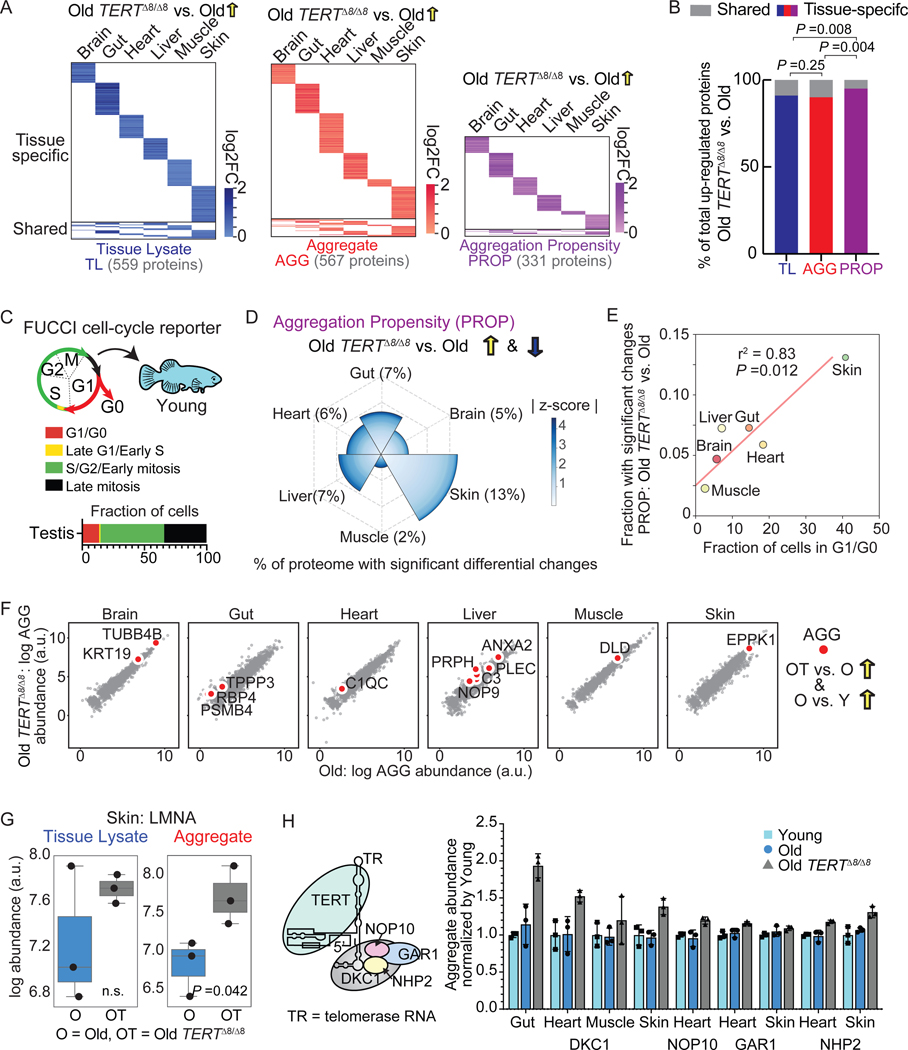
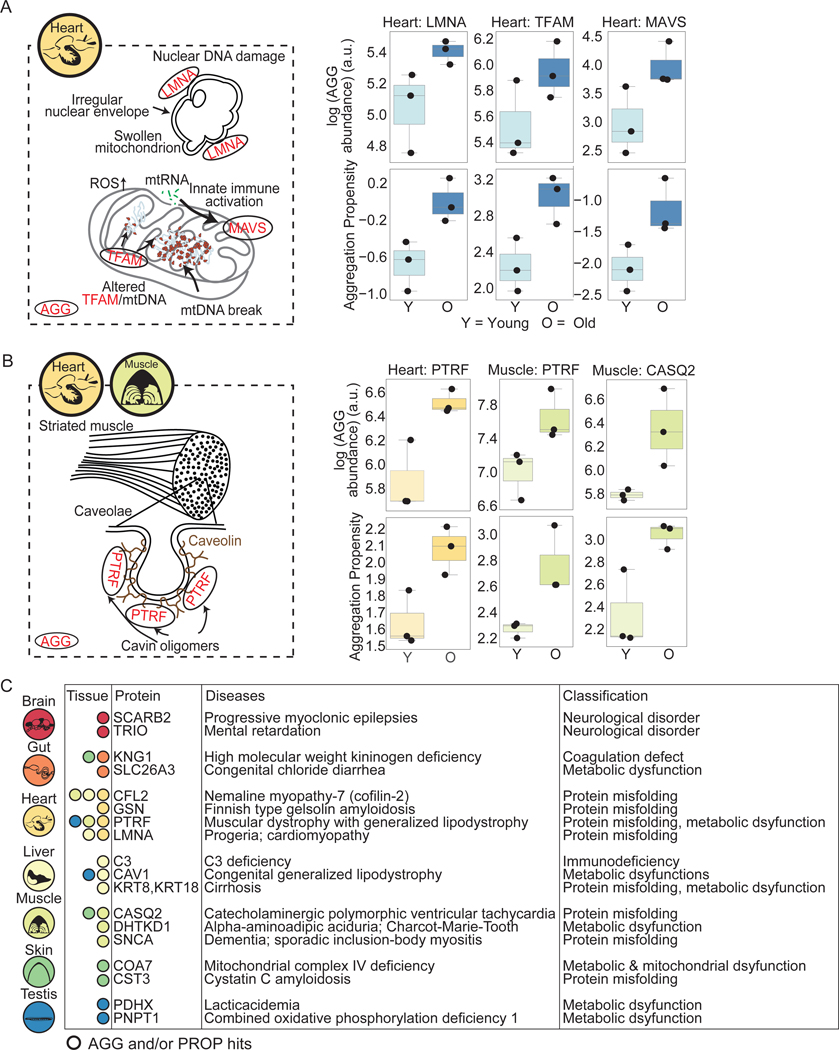
Protein aggregation is a hallmark of age-related neurodegeneration. Yet, aggregation during normal aging and in tissues other than the brain is poorly understood. Here, we leverage the African turquoise killifish to systematically profile protein aggregates in seven tissues of an aging vertebrate. Age-dependent aggregation is strikingly tissue specific and not simply driven by protein expression differences. Experimental interrogation in killifish and yeast, combined with machine learning, indicates that this specificity is linked to protein-autonomous biophysical features and tissue-selective alterations in protein quality control. Co-aggregation of protein quality control machinery during aging may further reduce proteostasis capacity, exacerbating aggregate burden. A segmental progeria model with accelerated aging in specific tissues exhibits selectively increased aggregation in these same tissues. Intriguingly, many age-related protein aggregates arise in wild-type proteins that, when mutated, drive human diseases. Our data chart a comprehensive landscape of protein aggregation during vertebrate aging and identify strong, tissue-specific associations with dysfunction and disease.
Keywords: aggregates; aging; multi-tissue; protein homeostasis; proteomics; systems biology.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Yang L, Cao Y, Zhao J, Fang Y, Liu N, and Zhang Y. (2019). Multidimensional Proteomics Identifies Declines in Protein Homeostasis and Mitochondria as Early Signals for Normal Aging and Age-associated Disease in Drosophila. Mol Cell Proteomics 18, 2078–2088. 10.1074/mcp.RA119.001621. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
