

Pancreatic beta-cell IL-22 receptor deficiency induces age-dependent dysregulation of insulin biosynthesis and systemic glucose homeostasis
- PMID: 38811550
- PMCID: PMC11137127
- DOI: 10.1038/s41467-024-48320-2
Pancreatic beta-cell IL-22 receptor deficiency induces age-dependent dysregulation of insulin biosynthesis and systemic glucose homeostasis
Abstract
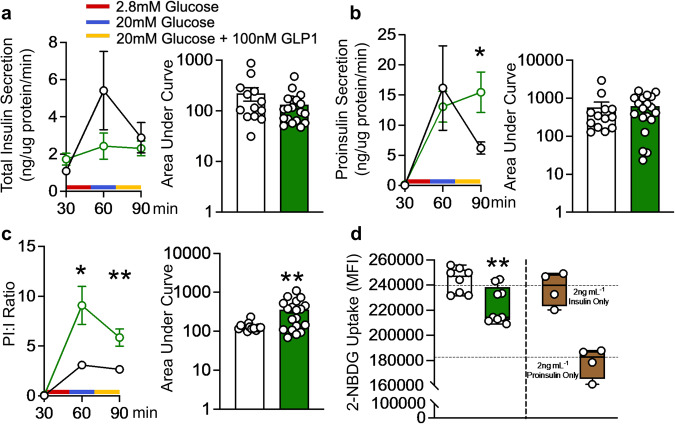
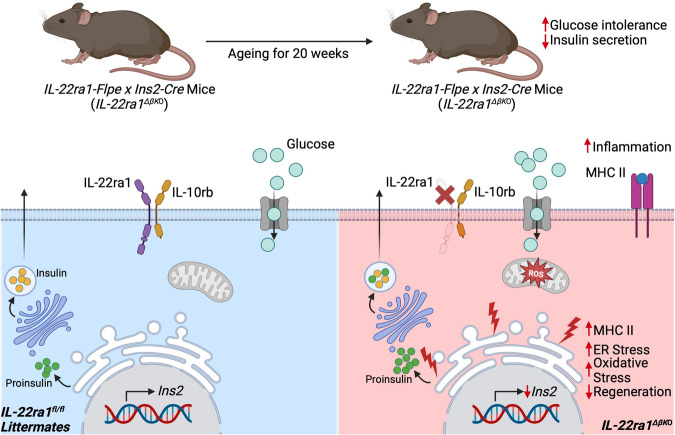
The IL-22RA1 receptor is highly expressed in the pancreas, and exogenous IL-22 has been shown to reduce endoplasmic reticulum and oxidative stress in human pancreatic islets and promote secretion of high-quality insulin from beta-cells. However, the endogenous role of IL-22RA1 signaling on these cells remains unclear. Here, we show that antibody neutralisation of IL-22RA1 in cultured human islets leads to impaired insulin quality and increased cellular stress. Through the generation of mice lacking IL-22ra1 specifically on pancreatic alpha- or beta-cells, we demonstrate that ablation of murine beta-cell IL-22ra1 leads to similar decreases in insulin secretion, quality and islet regeneration, whilst increasing islet cellular stress, inflammation and MHC II expression. These changes in insulin secretion led to impaired glucose tolerance, a finding more pronounced in female animals compared to males. Our findings attribute a regulatory role for endogenous pancreatic beta-cell IL-22ra1 in insulin secretion, islet regeneration, inflammation/cellular stress and appropriate systemic metabolic regulation.
© 2024. The Author(s).
Conflict of interest statement
S.Z.H, M.A.M and J.B.P are inventors on a patent relating to IL-22 use in metabolic disease. The remaining authors declare no other competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
