

The debranching enzyme Dbr1 regulates lariat turnover and intron splicing
- PMID: 38816363
- PMCID: PMC11139901
- DOI: 10.1038/s41467-024-48696-1
The debranching enzyme Dbr1 regulates lariat turnover and intron splicing
Abstract
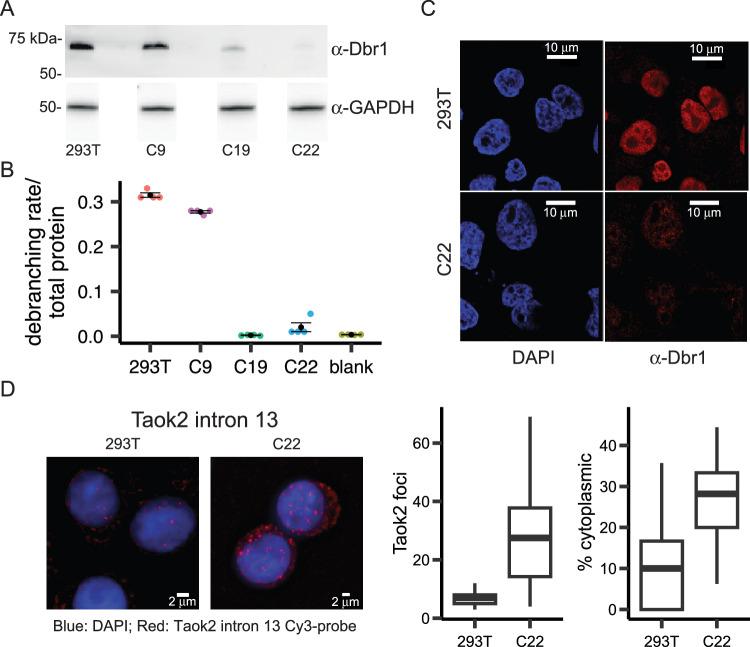
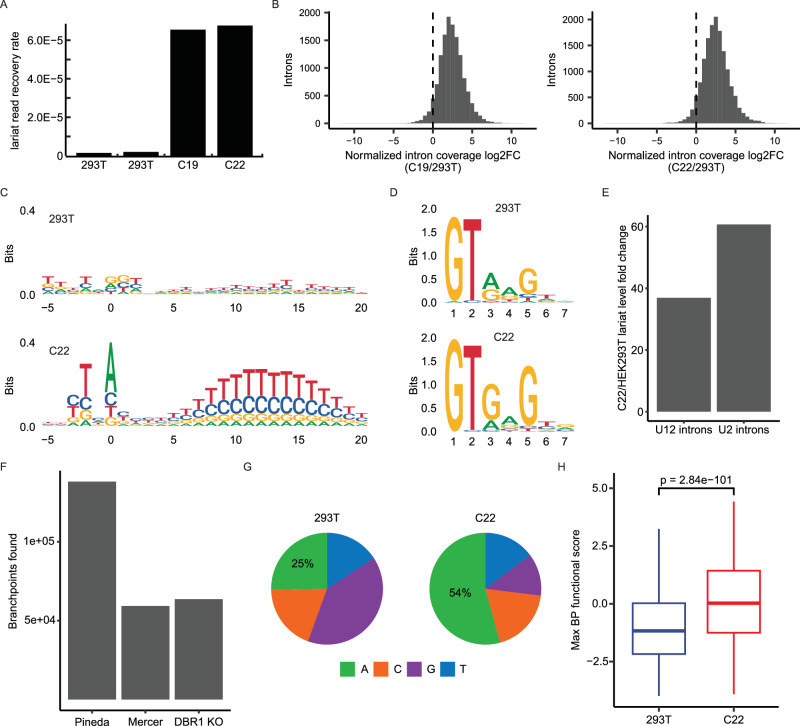
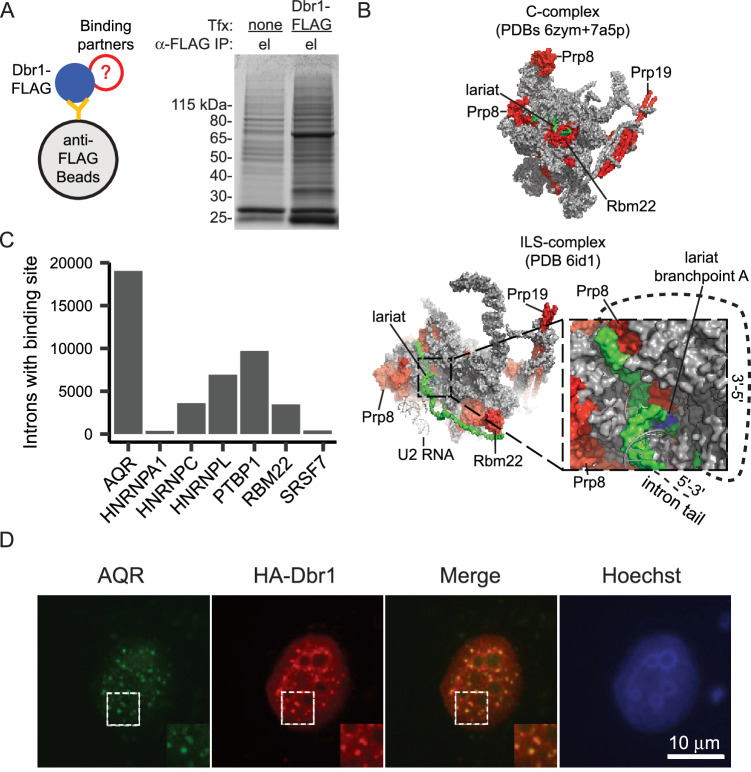
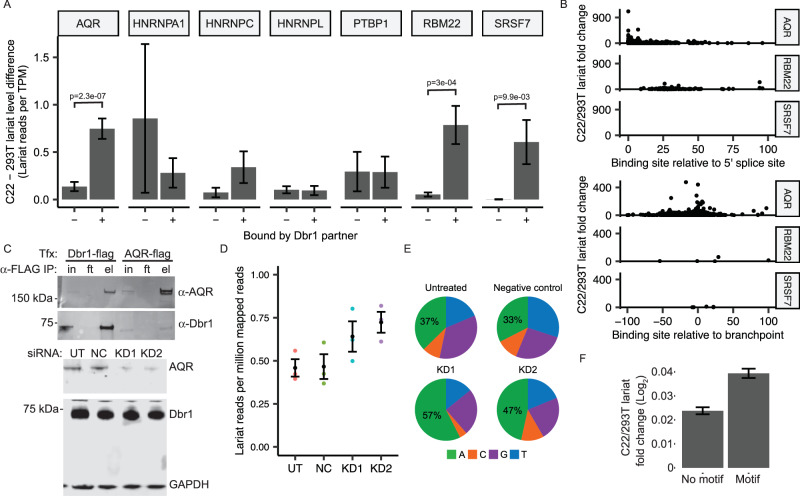
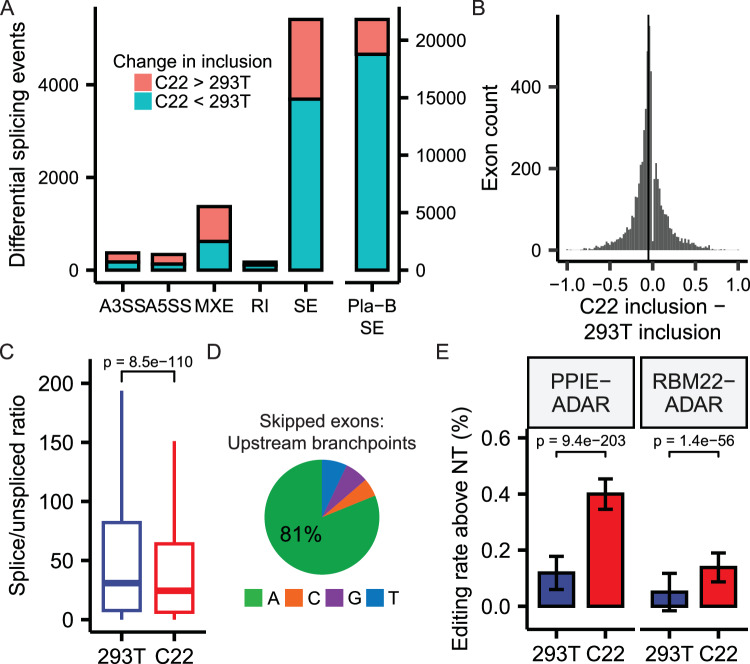
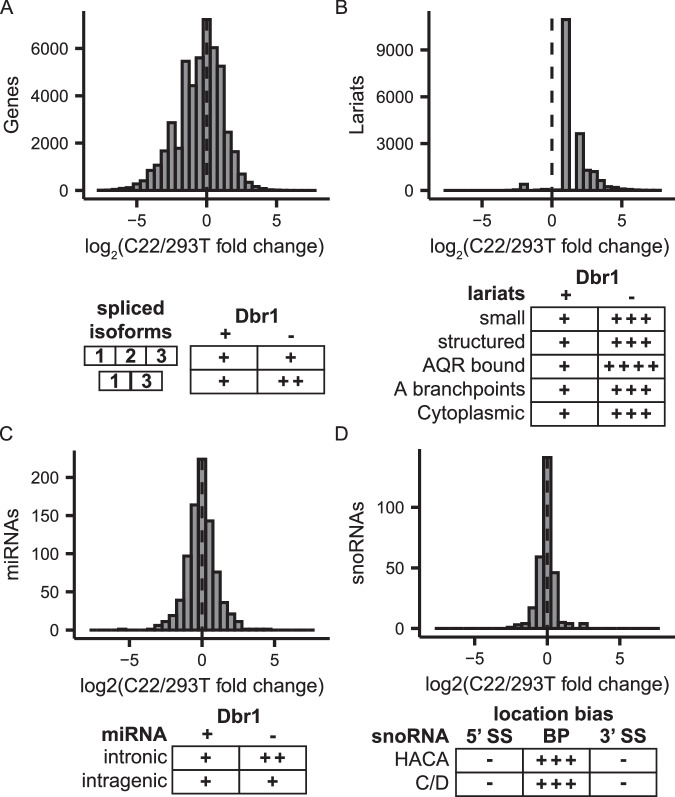
The majority of genic transcription is intronic. Introns are removed by splicing as branched lariat RNAs which require rapid recycling. The branch site is recognized during splicing catalysis and later debranched by Dbr1 in the rate-limiting step of lariat turnover. Through generation of a viable DBR1 knockout cell line, we find the predominantly nuclear Dbr1 enzyme to encode the sole debranching activity in human cells. Dbr1 preferentially debranches substrates that contain canonical U2 binding motifs, suggesting that branchsites discovered through sequencing do not necessarily represent those favored by the spliceosome. We find that Dbr1 also exhibits specificity for particular 5' splice site sequences. We identify Dbr1 interactors through co-immunoprecipitation mass spectrometry. We present a mechanistic model for Dbr1 recruitment to the branchpoint through the intron-binding protein AQR. In addition to a 20-fold increase in lariats, Dbr1 depletion increases exon skipping. Using ADAR fusions to timestamp lariats, we demonstrate a defect in spliceosome recycling. In the absence of Dbr1, spliceosomal components remain associated with the lariat for a longer period of time. As splicing is co-transcriptional, slower recycling increases the likelihood that downstream exons will be available for exon skipping.
© 2024. The Author(s).
Conflict of interest statement
We disclose W.G.F. as the founder of Walah Scientific and member of the scientific advisory board for Remix Therapeutics. All other authors declare no competing interests.
Figures
Update of
-
The debranching enzyme Dbr1 regulates lariat turnover and intron splicing.Res Sq [Preprint]. 2023 Jun 13:rs.3.rs-2931976. doi: 10.21203/rs.3.rs-2931976/v1. Res Sq. 2023. Update in: Nat Commun. 2024 May 30;15(1):4617. doi: 10.1038/s41467-024-48696-1. PMID: 37398028 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
- F31CA254143/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- R01 GM105681/GM/NIGMS NIH HHS/United States
- F31 CA254143/CA/NCI NIH HHS/United States
- RSG1815601DMC/American Cancer Society (American Cancer Society, Inc.)
- R01 CA193318/CA/NCI NIH HHS/United States
- S10 RR025111/RR/NCRR NIH HHS/United States
- P01 CA092584/CA/NCI NIH HHS/United States
- R01GM127472/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- P01CA092584/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- R01CA227001/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
- R01 GM127472/GM/NIGMS NIH HHS/United States
- R01GM105681/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R01 CA227001/CA/NCI NIH HHS/United States
- P30 CA091842/CA/NCI NIH HHS/United States
- R01CA193318/U.S. Department of Health & Human Services | NIH | National Cancer Institute (NCI)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
